Pain, Anxiety- and Depression-Like Behaviors in Alcohol-Preferring and -NonPreferring Rats
A B S T R A C T
Introduction: Psychiatric disorders such as anxiety, hyperalgesia, and depression have been associated with excessive alcohol drinking, but the neuronal mechanisms involved are only partially understood. Alcoholism more often occurs in individuals with a family history, indicating that genes may play a critical role. Chronic alcohol exposure alters calcium/calmodulin-dependent protein kinase II (CaMKII) signaling in the lateral habenula (LHb), and the LHb is implicated in mediating aversive behaviors, including those related to alcohol. We compared the CaMKII signaling in the LHb and the aberrant behaviors in the selectively bred alcohol-preferring (P) and alcohol-non-preferring (NP) lines of rats.
Materials and Methods: The responses to mechanical (Von Frey) and thermal (Hargreaves) nociception tests, anxiety- (elevated plus maze, Marble burying) and depressive-like behaviors (forced swimming) were examined in the alcohol-naïve P and NP rats, as well as in P rats after 4-8 weeks of alcohol consumption; their LHb tissues were also collected for Western blot analysis of CaMKII expression.
Results: Compared to NP rats, the P rats had a higher sensitivity to mechanical stimuli, and displayed depressive- and anxiety-like behaviors, as well as a higher level of CaMKII in the LHb. Alcohol consumption alleviated all these behaviors, except for anxiety, and decreased CaMKII levels in the LHb of P rats.
Conclusions: The results show that selective breeding for different oral alcohol preference has produced differences in nociception, anxiety, and depression, as well as CaMKII expression in the LHb of P and NP rats. P rats may deal with pain and depression by self-medicating with alcohol.
Keywords
Nociception, elevated plus maze, Hargreaves, Von Frey, marble burying, CaMKII
Introduction
Excessive repeated alcohol consumption is associated with certain psychiatric disorders such as anxiety, depression, and pain; however, the neuronal mechanisms involved are only partially understood. One possible mechanism is that these aberrant behaviors and alcohol use disorder (AUD) may share a common etiology, such as overlapping genetic risk factors [1, 2]. The sets of selectively bred alcohol-preferring- and alcohol-non-preferring lines, such as the ALKO alcohol/non-alcohol (AA/ANA) lines the Sardinian alcohol-preferring/alcohol-non-preferring (sP/sNP) lines, and the Indiana University alcohol-preferring/alcohol-non-preferring (P/NP) lines, may provide a useful platform to determine potential relationships between a predisposition toward high alcohol intake, psychiatric disorders and chronic pain [3-9].
Several studies have indicated a positive association between high alcohol intake, anxiety- and depressive-like behaviors. Alcohol-naïve sP rats have a higher anxiety level than sNP rats [10-12]. However, the relationship between alcohol intake and anxiety levels in P and NP rats remains controversial. While several studies reported a higher anxiety level in P rats than NP rats, other studies found no difference or even a lower anxiety level in P rats [8, 13-15]. Similarly, while alcohol-naïve sP and AA rats have shown a higher stress level than sNP and ANA rats, respectively [15, 16]. On the contrary, it was also reported that the P rats show less stress than the NP rats [17]. While growing evidence suggests that increased pain sensitivity might contribute to excessive drinking, it is unknown whether alcohol consumption is associated with pain sensitivity in P and NP lines of rats [18-20].
Recently, the lateral habenula (LHb) has emerged as a key brain region in the pathophysiology of depression, pain, and aversion [21-26]. The calcium/calmodulin-dependent protein kinase II (CaMKII) is a calcium-activated kinase enzyme and an essential mediator of various neuronal adaptive processes such as memory, addiction and pain [27-31]. Increased LHb CaMKII level has been associated with depressive-like behaviors, and inhibition of LHb CaMKII suppresses depressive-like behaviors and alcohol intake [32-34]. However, whether the P and NP rats inherently differ in the LHb CaMKII expression level is unknown.
In this study, we tested the hypothesis that alcohol-preferring rats could have inherently higher levels of CaMKII in the LHb and be more “anxious” and “depressed” with higher pain sensitivity than their non-preferring counterparts.
Materials and Methods
I Animals and Housing
Indiana University School of Medicine Animal Research Center (Indianapolis, IN, USA) provided the selectively bred male alcohol-preferring (P) and alcohol-non-preferring (NP) rats. These were shipped to Rutgers University at approximately 60 days of age and underwent routine quarantine procedures for at least eight weeks upon arrival. They were then single-housed in ventilated Plexiglas cages in a climate-controlled room (20-22℃) with a 12-h light/dark cycle (light-off at 11:00 a.m.). Food and water were available ad libitum. The Institutional Animal Care and Use Committees of Rutgers approved all studies by the guidelines of the National Institutes of Health (NIH) for the Care and Use of Laboratory Animals. All of the P and NP rats were kept in the same room of the animal center. The alcohol naïve rats had two bottles of water, while the ethanol-exposed P (PE) rats enrolled in the intermittent access 20% ethanol two-bottle choice drinking procedure. We changed the water of the naïve rats at the same time when we changed the ethanol and water for the PE rats. Eight alcohol naïve P and six NP rats were subjected to behavioral tests starting at the age of 4 months while simultaneously, 12 P rats were subjected to ethanol consumption (see below for details).
II Intermittent Access 20% Ethanol Two-Bottle Choice Drinking Procedure (IA2BC)
P rats (n=12) were trained to drink alcohol under the IA2BC paradigm [34-37]. Briefly, rats were given 24-hour concurrent access to two bottles, one with 20% ethanol (v/v) and another with water, starting at 11:00 a.m. on Monday. After 24 hours, the ethanol bottle was replaced with a second water bottle, which was available for the next 24 hours. This pattern was repeated on Wednesdays and Fridays. On all other days, the rats had unlimited access to two bottles of water. In each ethanol drinking session, the placement of the ethanol bottle was alternated to control for side preferences. The amount of ethanol or water consumed was determined by weighing the bottles before and after 24-hours of access. The body weight of each rat was measured weekly to monitor the health and calculate the grams of ethanol intake per kilogram of body weight. The following formula derived the preferred ratio of ethanol intake: Preference ratio (%) = ethanol solution intake (ml)/total fluid intake (ml of ethanol solution + ml of water). Behavioral tests were conducted on alcohol-naïve P rats, ethanol-exposed P rats (after 4 weeks of ethanol drinking), as well as on alcohol-naive NP rats.
III Blood Ethanol Concentration Measurement
Blood samples (around 0.4 ml) of P rats (n = 8) were collected from the lateral tail vein immediately after the 2-hour, and the 24-hour access to ethanol, as well as at 24 h withdrawal from the last ethanol session at the end of the 8-week IA2BC drinking schedule. Blood ethanol concentration was measured as described previously [9]. Briefly, the blood samples were centrifuged at 4℃ for 15 minutes at 8000 rpm, and 10 µl plasma from each blood sample was analyzed using the nicotinamide adenine dinucleotide-alcohol dehydrogenase (NAD-ADH) enzyme spectrophotometric method [38].
IV Pain Threshold Tests
The up-down testing paradigm was applied to evaluate the paw withdrawal threshold (PWT) in response to mechanical stimuli, as described previously [39]. In this process, the rats were placed in a Plexiglas chamber on an elevated mesh screen and allowed to acclimate for at least 15 minutes. A researcher who was blinded to the line of rats measured the PWT. A series of von Frey hairs in log increments of force were applied perpendicularly to the plantar surface of the hind paw for 3 seconds. A sharp withdrawal of the hind paw indicated a positive response. When a positive response occurred, a smaller von Frey hair would be used; while if a negative response occurred, a larger one would be used. The PWT was determined by converting the pattern of positive and negative responses to the von Frey filament stimulation with the formula provided by Dixon [40].
Paw withdrawal latencies (PWL) to thermal stimuli were measured with an Analgesia Meter (Model 336, IITC Life Science Instruments, Woodland Hills, CA, USA) as described previously [39]. Briefly, each rat was placed in a Plexiglas chamber on a glass plate located above a lightbox. After a 15-minute acclimation to the environment, these rats were subjected to radiant heat, which was applied by aiming a light beam to the middle of the plantar surface of the hind paw. When the rat lifted its paw in response to the heat, the light beam was turned off. The PWL, measured in seconds, was defined as the average length of time between the start of the light beam and the foot-lift. Each measure was repeated five times, allowing 5 minutes between successive tests to avoid sensitization to the stimulus. A cutoff time of 20 seconds was used to prevent paw tissue damage.
V Measurement of Anxiety-Like Behaviors
The anxiety-like behaviors were evaluated with an elevated plus maze test and a marble burying test, as described previously [25, 41].
VI Elevated Plus Maze Test (EPM)
The maze consisted of four-arms (two open without walls and two enclosed by 30 cm high walls) perpendicular to a central platform 50 cm above the floor and was placed in an isolated silent and dimly lit room (illumination of 100-140 lux). The time spent in the open arms, the number of entries into open arms, and the total distance traveled were recorded using Smart 3.0 (Pan lab Harvard Apparatus, Barcelona, Spain).
VII Marbles Burying Test (MBT)
In a standard rat cage, twenty 1.5 cm glass marbles were evenly spaced in five rows of four on a 4-5 cm layer of corncob bedding materials. A rat was placed in the test cage and the number of marbles buried in the bedding (to 2/3 their depth) after 30 min was counted.
VIII Forced Swimming Test (FST)
To measure general depressive-like behaviors, we adapted a commonly used modified FST in a transparent plastic tube (diameter = 24.5 cm, height = 51 cm) filled to 30 cm with water at 23-25 °C [34, 42]. In the first session of the test, each rat was placed in the tank of water for 15 min. After 24 h, the rat was placed into the tank again under the same conditions for another 5 min. Immobility was defined when the rat, floating in the water without struggling, only made small and necessary movements to keep its head above the water.
IX Western Blot Analysis
After behavioral tests, the rats were sacrificed for Western blot analysis. Rats were deeply anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and then transcardially perfused with saline. Tissue containing the LHb of both hemispheres was harvested and homogenized on ice in radio-immunoprecipitation assay buffer (Sigma-Aldrich, St. Louis, MO). Proteins were quantified using the Bradford assay (Bio-Rad Laboratories). Using the Trans-Blot Turbo Transfer System (Bio-Rad Laboratories), ten μg per sample was loaded on an SDS-PAGE gel and transferred to a polyvinylidene difluoride (PVDF) membrane. The protein level of CaMKII and GAPDH in the LHb were detected by anti-CaMKII (1:2000, Thermo Scientific) and anti-GAPDH antibody (1:2500, Sigma-Aldrich), respectively. We quantified the western blots in 2 steps. First, protein loading and relative protein expression were compared with GAPDH levels in each lane by quantitatively analyzing the optical densities of bands using ImageJ version 1.38 (NIH, Bethesda, MD). Then, we presented changes in protein levels of alcohol-naïve P rats and ethanol-exposed P rats as a percentage of those in the NP group (set to 100).
X Statistical Analysis
In order to extract significant discrepancy, one-way ANOVA was conducted on the behavior test data and the optical density from western blot analysis among the three groups. If the data were normally distributed, the post hoc comparisons were assessed using Tukey’s test. Otherwise, Dunn's multiple range test was used for post hoc comparisons. In all the analyses, p < 0.05 were accepted as being statistically significant.
Results
I Ethanol Intake and Preference in P Rats
P rats consumed a high amount of ethanol and had a high preference for ethanol at the first drinking session (11.30 ± 2.24 g/kg/24 h and 70.12 ± 3.20% preference, n=12). Over the 11 drinking sessions in the IA2BC paradigm, both ethanol intake (11.05 ± 0.51 g/kg/24 h) and preference (74.04 ± 1.52%) were not significantly changed (ethanol intake, p = 0.260; ethanol preference, p = 0.244, Figures 1A & 1B). Blood ethanol concentrations (BECs) measured immediately after the 2-hour, and 24-hour access to ethanol, as well as at 24-hour withdrawal from ethanol, were respectively 66.4 ± 6.6 (with 71% was above 60 mg/dl), 45.5 ± 5.1 mg/dl and 29.3 ± 2.7 mg/dl.
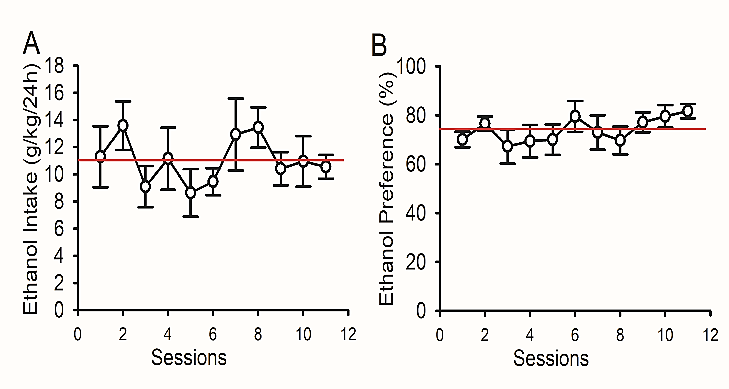
Figure 1: Ethanol intake and preference in P rats. P rats (n=12) were trained to drink ethanol under the intermittent access to 20% ethanol in a two-bottle choice drinking paradigm.
II Mechanical Nociception
Alcohol-naïve P rats exhibited higher sensitivity to mechanical stimuli as reflected by the lower PWT in response to the Von Frey fibers in comparison to the alcohol-naïve NP rats (P vs. NP, p < 0.001, Figure 2A). Four-week ethanol drinking in the IA2BC paradigm significantly increased the PWT (P vs. ethanol-exposed P, p = 0.009, Figure 2A), without altering the PWL in response to thermal stimuli. We found no significant difference in the PWL among P, NP and ethanol-exposed P rats (all p > 0.05, Figure 2B).
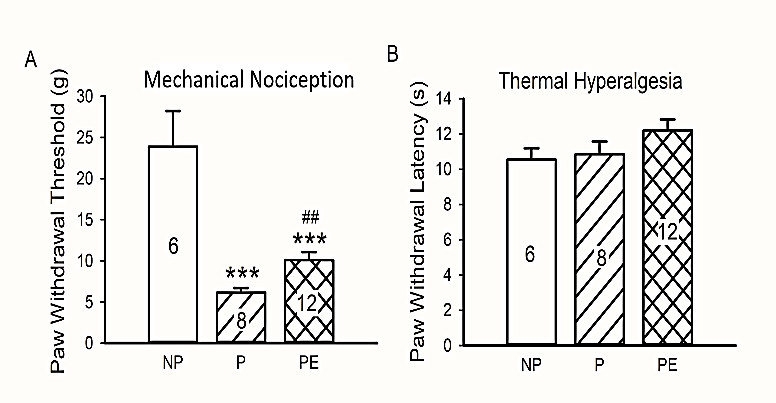
Figure 2: Paw withdrawal responses to thermal and mechanical stimuli in NP, P and P rats exposed to ethanol (PE). A) Paw withdrawal threshold (PWT) response to mechanical stimuli. B) Paw withdrawal latency (PWL) response to thermal stimuli. ***p < 0.001 vs. NP, ##p < 0.01 vs. P. Animal numbers are indicated.
III Anxiety-Like Behaviors
Anxiety-like behaviors were assessed using elevated plus maze (EMP) and marble burring test (MBT). P rats exhibited a higher anxiety level as reflected by spending a shorter time in the open arms (P vs. NP, p < 0.001, Figure 3A), entered the open arms less frequently (P vs. NP, p < 0.05, Figure 3B), and traveled a shorter distance (P vs. NP, p < 0.001, Figure 3C) in comparison to the NP rats.
Four-week ethanol consumption dramatically increased the total distance traveled by the P rats (P vs. ethanol-exposed P, p = 0.007, Figure 3C), without altering the time in the open arms (P vs. ethanol-exposed P, p = 0.9, Figure 3A) or the entries to the open arms (P vs. ethanol-exposed P, p = 0.8, Figure 3C). Compared with the NP rats, P rats buried more marbles in the MBT (P vs. NP, p < 0.05, Figure 3D), which was also not significantly changed by ethanol consumption (P vs. ethanol-exposed P, p > 0.05, Figure 3D).
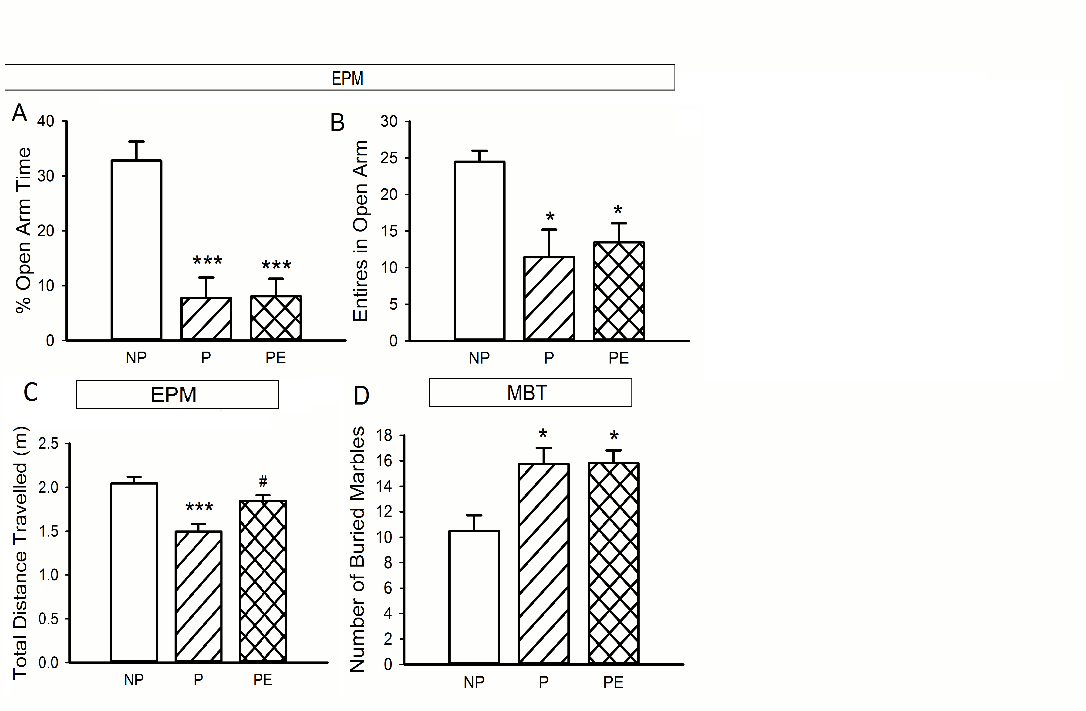
Figure 3: Anxiety-like behavior in P, NP, and P rats exposed to ethanol. A) Percent of time in the open arms in the EPM. Compared with the NP rats, alcohol-naïve P rats or P rats after chronic ethanol consumption spent a significantly shorter time in the open arms and B) entered less time to the open arms in the EPM. C) P rats traveled a shorter distance in the open arms than NP did. Ethanol exposure increased the total distance traveled by P rats. D) Marbles buried in MBT. P rats in naïve and after ethanol exposure buried more marbles than NP did. *p <0.05, ***p < 0.001 vs. NP; #p < 0.05 vs. P.
IV Depression-Like Behavior
Depression-like behavior was assessed using the forced swimming test (FST). Compared to NP rats, P rats exhibited higher sensitivity to stress as reflected by the longer immobility time (P vs. NP, p < 0.05, Figure 4A), which was reversed by ethanol consumption (P vs. ethanol-exposed P, p < 0.05, Figure 4).
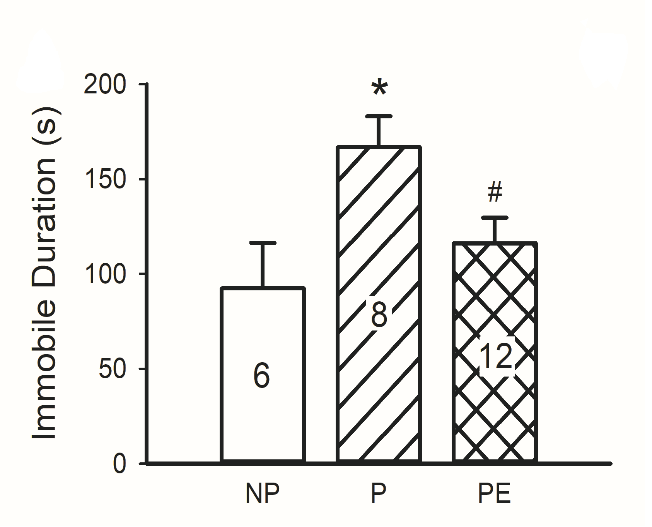
Figure 4: Depressive-like behaviour in NP, P and P rats exposed to ethanol (PE). Immobile duration in the Forced Swimming Test (FST). P rats displayed longer immobility duration in the FST than NP rats did. Ethanol exposure reduced immobility duration in P rats. *p < 0.05, P vs. NP; #p < 0.05 PE vs. P. Animal numbers are indicated.
V CaMKII Levels in the LHb
Compared to NP rats, the P rats had a significantly higher level of CaMKII protein expression level in the LHb (P vs. NP, p < 0.001, Figures 5A & 5B), which was reversed by ethanol consumption (P rats vs. ethanol-exposed P rats, p < 0.05, Figures 5A & 5B).
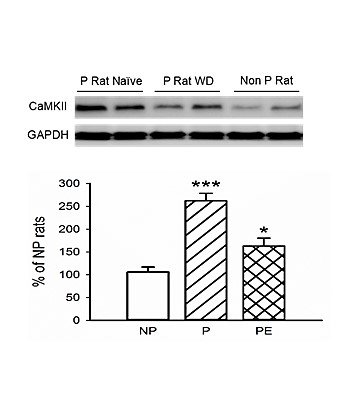
Figure 5: LHb CaMKII levels in P, NP and P rats exposed to ethanol. A) Representative Western blot image of CaMKII. B) Analysis to quantify expression levels of CaMKII in the naïve P rats (P), P rats exposed to ethanol (PE) and non-P rats (NP). P rats displayed a higher level of CaMKII in the LHb than NP rats. Ethanol exposure notably attenuated CaMKII level in the LHb in P rats. ***p < 0.001 P vs. NP, *p < 0.05 vs. P. n = 3/arm.
Discussion
Compared to NP rats, P rats displayed higher anxiety-like and depressive-like behaviors, exhibited greater mechanical nociception, and had a higher level of CaMKII in the LHb, all of which, except the anxiety level, were alleviated by 4-8 weeks of ethanol consumption.
We and others had previously reported that Long-Evans and Sprague-Dawley rats significantly escalate the intake of and the preference for ethanol in the IA2BC procedure [34-36, 43, 44]. In the current study, P rats drank a high amount of ethanol and had a high preference in the first drinking session, consistent with previous reports [7, 13]. Remarkably, ethanol intake and preference were not changed over the course of 11 drinking sessions in the IA2BC paradigm. The mechanisms underlying the lack of further increase are unclear. We speculate that the high amount of initial intake may be a major factor.
There is increasing interest in the relationship between AUD and pain. Acute alcohol has an analgesic effect [45]. However, chronic alcohol use can produce paradoxical hyperalgesia [18, 46]. Alcoholics are usually more sensitive to painful stimulation than non-alcoholic individuals [47]. Even people who only have a family history of alcoholism but no drinking problems themselves are more sensitive to pain than those who do not have a family history of alcohol addiction [48]. In the current study, we observed that the alcohol-naïve P rats were more sensitive to mechanical stimuli than the alcohol-naive NP rats. Higher pain sensitivity may contribute to alcohol abuse and addiction [49]. We previously reported that hyperalgesia during alcohol withdrawal could promote relapse-like drinking [25, 39]. Gatch and Harbans reported that the acute and chronic administration of ethanol produced antinociception and that tolerance developed with chronic administration. Moreover, withdrawal-induced hyperalgesia was reversed by relapse [18, 19].
There was no difference in thermal nociception between the alcohol-naïve P and alcohol-naïve NP rats, consistent with previous reports [13, 50]. Ethanol administration did not alter thermal nociception in P rats, which is consistent with a previous report [51]. Conversely, the strain of rats with high alcohol consumption (AA rats) displays thermal hyperalgesia relative to their low consumption parallel strain (ANA rats) [52]. The selected High Alcohol Drinking (HAD1) and Low Alcohol Drinking (LAD1) lines of rats also have no difference in the paw withdrawal latency, but the LAD1 rats have a shorter latency in the tail flick assay [50].
In contrast to the lack of difference in thermal nociception, the alcohol-naïve P rats had a lower mechanical pain threshold than the alcohol-naïve NP rats. A previous study suggests that genetic variation in pain responses might depend on the type of pain studied and suggested dissociating mechanical and thermal nociceptive modalities [53]. Thermal and mechanical pain are served by distinct primary afferent fiber types, which are located in different spinal cord laminae and distinct brain regions [54-56]. Our data suggest that genetic differences might lead to the variance in response to mechanical nociception, but not the thermal hyperalgesia, between P and NP rats.
Anxiety more often occurs in alcoholics [25, 41]. Earlier studies comparing anxiety levels between P and NP rats are controversial. There are reports that the P rats have a higher anxiety level than the NP rats [8, 57, 58]. Others reported that P rats exhibit less anxiogenic behaviors [14]. Also, there are studies that found no differences between P and NP rats [12, 15]. We found P rats spent a shorter time in, and entered less into, the open arms than NP rats in the EPM, suggesting that P rats had a higher anxiety level. This idea is further supported by the result showing that P rats buried more marbles in MBT. Intriguingly, ethanol experience did not significantly alter the anxiety level of P rats. Additionally, we noticed the P rats traveled a significantly shorter distance in the EPM than the NP rats. Therefore, we analyzed the percent of time and entries in the open arms to avoid the possible influence of the different mobility.
Depression has been linked to alcohol abuse and relapse drinking [59, 60]. In this study, we found that compared to the alcohol-naïve NP rats, the alcohol-naïve P rats exhibited longer immobility in the FST, which was reversed by alcohol consumption. These results are similar to the difference between sP and sNP, as well as AA and ANA rats, but conflicting with the results of P/NP studies [15, 60-62]. The reason causing the difference is unclear, but we propose that the conditions in the FST experiments might matter. In our study as well as that of Ciccocioppo et al., FST was conducted with a 30 cm water depth, where rats could touch the bottom of the syringe with their tail but could not support themselves with their hindlimbs [16]. Conversely, in Godfrey and Ehlers’ studies, FST was conducted with a water depth of 35 cm or 48 cm, respectively, where the rat was not able to touch the bottom [17, 62]. Future studies are needed to confirm this hypothesis.
We previously reported that CaMKII in the LHb of Long-Evans rats was increased after chronic alcohol drinking and that inhibition of LHb CaMKII activity reduced ethanol consumption, and depressive-like behaviors [34]. In the current study, we found that the CaMKII level in the LHb of alcohol-naïve P rats was higher than the NP rats. Therefore, we speculate that the increased CaMKII expression in the LHb might be associated with the inherently higher alcohol preference of the P rats. After chronic ethanol consumption, the CaMKII concentration in the LHb of P rats was significantly decreased to the level similar to the NP rats. Interestingly, chronic alcohol drinking increased and decreased CaMKII in the LHb of Long-Evans rats and P rats, respectively. Thus, the pathological effects of chronic alcohol drinking might vary on rats with different genetic backgrounds. Moreover, the different alterations of the CaMKII concentration between the P line and Long-Evans line of rats may also result from the dose-effect, because of the gap in the amount of alcohol consumed. CaMKII has been linked with pain hypersensitivity [30, 31]. The higher CaMKII levels in the LHb of P rats may also be associated with their higher sensitivity to mechanical stimuli.
The present study indicated there were varieties in behaviors and CaMKII expression among P and NP rats, as well as the P rats exposed to ethanol, but we did not include the NP rats with alcohol exposure in our study. The performs of NP rats exposed to ethanol could be investigated in a future study, which might improve our results. As the genetic background may play an important role in these varieties, the Wistar rats, which line the P and NP rats were developed from, should also be considered as a control to draw a clearer pattern and confirm our current results.
In conclusion, the alcohol-preferring rats displayed higher sensitivity to mechanical nociception and had higher levels of anxiety and depression than the alcohol-non-preferring rats. Alcohol drinking alleviated the mechanical nociception and depressive-like behaviors, however not the anxiety-like behaviors in P rats. Moreover, the levels of CaMKII protein in the LHb of P rats were higher than that of NP rats, which were decreased after chronic alcohol consumption. The inborn alcohol preference of the P rats and their behavioral and neurobiological features may have an association with their genetic background. Chronic alcohol drinking might cause changes in mechanical nociception and depression by altering the epigenetic regulation on LHb CaMKII expression. Thus, we propose the CaMKII as one of the possible candidates involved in alcohol dependence, mechanical nociception, and depressive-like behaviors. Alcohol may be implicated in reducing pain and depression by modulating the LHb CaMKII levels of P rats.
Funding
This work was supported by the National Institute of Health Grants AA021657 and AA022292.
Conflicts of Interest
None.
Acknowledgments
The authors acknowledge Indiana University for providing the P- and Non-P rats, and Manideepika Katta for reading the manuscript.
Article Info
Article Type
Research ArticlePublication history
Received: Fri 24, Apr 2020Accepted: Wed 06, May 2020
Published: Tue 12, May 2020
Copyright
© 2023 Jiang-Hong Ye. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.NNB.2020.02.05
Figures & Tables
References
- Gilpin NW, Koob GF (2008) Neurobiology of alcohol dependence: focus on motivational mechanisms. Alcohol Res Health 31: 185-195. [Crossref]
- Becker HC (2008) Alcohol dependence, withdrawal, and relapse. Alcohol Res Health 31: 348-361. [Crossref]
- Sommer W, Hyytia P, Kiianmaa K (2006) The alcohol-preferring AA and alcohol-avoiding ANA rats: neurobiology of the regulation of alcohol drinking. Addict Biol 11: 289-309. [Crossref]
- Eriksson K (1968) Genetic selection for voluntary alcohol consumption in the albino rat. Science 159: 739-741. [Crossref]
- Colombo G, Lobina C, Carai MAM, Gessa GL (2006) Phenotypic characterization of genetically selected Sardinian alcohol-preferring (sP) and -non-preferring (sNP) rats. Addict Biol 11: 324-338.
- Lobina C, Agabio R, Diaz G, Fa M, Fadda F et al. (1997) Constant absolute ethanol intake by Sardinian alcohol-preferring rats independent of ethanol concentrations. Alcohol Alcohol 32: 19-22. [Crossref]
- Lumeng L, Hawkins T, Li TK (1977) New strains of rats with alcohol preference and nonpreference, in Alcohol and Aldehyde Metabolizing Systems: Intermediary Metabolism and Neurochemistry. Elsevier 537-544.
- Stewart RB, Gatto GJ, Lumeng L, Li TK, Murphy JM (1993) Comparison of alcohol-preferring (P) and nonpreferring (NP) rats on tests of anxiety and for the anxiolytic effects of ethanol. Alcohol 10: 1-10. [Crossref]
- Bell RL, Sable HJ, Colombo G, Hyytia P, Rodd ZA et al. (2012) Animal models for medications development targeting alcohol abuse using selectively bred rat lines: neurobiological and pharmacological validity. Pharmacol Biochem Behav 103: 119-155. [Crossref]
- Colombo G, Agabio R, Lobina C, Reali R, Zocchi A et al. (1995) Sardinian alcohol-preferring rats: a genetic animal model of anxiety. Physiol Behav 57: 1181-1185. [Crossref]
- Colombo G, Agabio R, Lobina C, Reali R, Melis F et al. (1995) Effects of the calcium channel antagonist darodipine on ethanol withdrawal in rats. Alcohol Alcohol 30: 125-131. [Crossref]
- Roman E, Stewart RB, Bertholomey ML, Jensen ML, Colombo G et al. (2012) Behavioral profiling of multiple pairs of rats selectively bred for high and low alcohol intake using the MCSF test. Addict Biol 17: 33-46. [Crossref]
- Kampov Polevoy AB, Matthews DB, Gause L, Morrow AL, Overstreet DH (2000) P rats develop physical dependence on alcohol via voluntary drinking: changes in seizure thresholds, anxiety, and patterns of alcohol drinking. Alcohol Clin Exp Res 24: 278-284. [Crossref]
- Overstreet DH (1997) Behavioral similarities and differences among alcohol-preferring and -nonpreferring rats: confirmation by factor analysis and extension to additional groups. Alcohol Clin Exp Res 21: 840-848.
- Viglinskaya IV, Overstreet DH, Kashevskaya OP, Badishtov BA, Kampov Polevoy AB et al. (1995) To drink or not to drink: tests of anxiety and immobility in alcohol-preferring and alcohol-nonpreferring rat strains. Physiol Behav 57: 937-941. [Crossref]
- Ciccocioppo R, Panocka I, Froldi R, Colombo G, Gessa GL et al. (1999) Antidepressant-like effect of ethanol revealed in the forced swimming test in Sardinian alcohol-preferring rats. Psychopharmacology (Berl) 144: 151-157. [Crossref]
- Godfrey CD, Froehlich JC, Stewart RB, Li TK, Murphy JM (1997) Comparison of rats selectively bred for high and low ethanol intake in a forced-swim-test model of depression: effects of desipramine. Physiol behav 62: 729-733. [Crossref]
- Gatch MB (2009) Ethanol withdrawal and hyperalgesia. Curr Drug Abuse Rev 2: 41-50. [Crossref]
- Gatch MB, Lal H (1999) Effects of ethanol and ethanol withdrawal on nociception in rats. Alcohol Clin Exp Res 23: 328-333. [Crossref]
- Jochum T, Boettger MK, Burkhardt C, Juckel G, Bär KJ (2010) Increased pain sensitivity in alcohol withdrawal syndrome. Eur J Pain 14: 713-718. [Crossref]
- Zhang CX, Zhang H, Xu HY, Li MX, Wang S (2013) The lateral habenula is a common target of cocaine and dexamethasone. Neurosci Lett 555: 12-17. [Crossref]
- Matsumoto M, Hikosaka O (2009) Representation of negative motivational value in the primate lateral habenula. Nat Neurosci 12: 77-84. [Crossref]
- Yang Y, Cui Y, Sang K, Dong Y, Ni Z et al. (2018) Ketamine blocks bursting in the lateral habenula to rapidly relieve depression. Nature 554: 317-322. [Crossref]
- Kang S, Li J, Zuo W, Chen P, Gregoe D et al. (2019) Downregulation of M-channels in lateral habenula mediates hyperalgesia during alcohol withdrawal in rats. Sci Rep 9: 2714.
- Gregor DM, Zuo W, Fu R, Bekker A, Ye JH (2019) Elevation of Transient Receptor Potential Vanilloid 1 Function in the Lateral Habenula Mediates Aversive Behaviors in Alcohol-withdrawn Rats. Anesthesiology 130: 592-608. [Crossref]
- Zuo W, Fu R, Hopf FW, Xie G, Krnjević K et al. (2017) Ethanol drives aversive conditioning through dopamine 1 receptor and glutamate receptor-mediated activation of lateral habenula neurons. Addict Biol 22: 103-116. [Crossref]
- Yamauchi T (2005) Neuronal Ca2+/calmodulin-dependent protein kinase ii—discovery, progress in a quarter of a century, and perspective: implication for learning and memory. Biol Pharm Bull 28: 1342-1354. [Crossref]
- Lee AM, Messing RO (2008) Protein kinases and addiction. Ann N Y Acad Sci 1141: 22-57. [Crossref]
- Müller CP, Quednow BB, Lourdusamy A, Kornhuber J, Schumann G et al. (2016) CaM Kinases: From Memories to Addiction. Trends Pharmacol Sci 37: 153-166. [Crossref]
- Luo F, Yang C, Chen Y, Shukla P, Tang L et al. (2008) Reversal of chronic inflammatory pain by acute inhibition of Ca2+/calmodulin-dependent protein kinase II. J Pharmacol Exp Ther 325: 267-275. [Crossref]
- Gwak YS, Hassler SE, Hulsebosch CE (2013) Reactive oxygen species contribute to neuropathic pain and locomotor dysfunction via activation of CamKII in remote segments following spinal cord contusion injury in rats. Pain 154: 1699-1708. [Crossref]
- Kim Y, Morath B, Hu C, Byrne LK, Sutor SL et al. (2016) Antidepressant actions of lateral habenula deep brain stimulation differentially correlate with CaMKII/GSK3/AMPK signaling locally and in the infralimbic cortex. Behav Brain Res 306: 170-177. [Crossref]
- Li K, Zhou T, Liao L, Yang Z, Wong C et al. (2013) βCaMKII in lateral habenula mediates core symptoms of depression. Science 341: 1016-1020. [Crossref]
- Li J, Kang S, Fu R, Wu L, Wu W et al. (2017) Inhibition of AMPA receptor and CaMKII activity in the lateral habenula reduces depressive-like behavior and alcohol intake in rats. Neuropharmacology 126: 108-120. [Crossref]
- Simms JA, Steensland P, Medina B, Abernathy KE, Chandler LJ et al. (2008) Intermittent access to 20% ethanol induces high ethanol consumption in Long–Evans and Wistar rats. Alcohol Clin Exp Res 32: 1816-1823. [Crossref]
- Li J, Zuo W, Fu R, Xie G, Kaur A et al. (2016) High Frequency Electrical Stimulation of Lateral Habenula Reduces Voluntary Ethanol Consumption in Rats. Int J Neuropsychopharmacol 19. [Crossref]
- Fu R, Zuo W, Gregor D, Li J, Grech D et al. (2016) Pharmacological Manipulation of the Rostromedial Tegmental Nucleus Changes Voluntary and Operant Ethanol Self-Administration in Rats. Alcohol Clin Exp Res 40: 572-582. [Crossref]
- Poklis A, Mackell MA (1982) Evaluation of a modified alcohol dehydrogenase assay for the determination of ethanol in blood. Clin Chem 28: 2125-2127. [Crossref]
- Fu R, Gregor D1, Peng Z1, Li J1, Bekker A et al. (2015) Chronic intermittent voluntary alcohol drinking induces hyperalgesia in Sprague-Dawley rats. Int J Physiol Pathophysiol Pharmacol 7: 136-144. [Crossref]
- Dixon WJ (1980) Efficient Analysis of Experimental Observations. Annu Rev Pharmacol Toxicol 20: 441-462. [Crossref]
- Kang S, Li J, Zuo W, Fu R, Gregor D et al. (2017) Ethanol Withdrawal Drives Anxiety-Related Behaviors by Reducing M-type Potassium Channel Activity in the Lateral Habenula. Neuropsychopharmacology 42: 1813-1824. [Crossref]
- Slattery DA, Uschold N, Magoni M, Bär J, Popoli M et al. (2012) Behavioural consequences of two chronic psychosocial stress paradigms: anxiety without depression. Psychoneuroendocrinology 37: 702-714. [Crossref]
- Carnicella S, Ron D, Barak S (2014) Intermittent ethanol access schedule in rats as a preclinical model of alcohol abuse. Alcohol 48: 243-252. [Crossref]
- Fu R, Chen X, Zuo W, Li J, Kang S et al. (2016) Ablation of mu opioid receptor-expressing GABA neurons in rostromedial tegmental nucleus increases ethanol consumption and regulates ethanol-related behaviors. Neuropharmacology 107: 58-67. [Crossref]
- Petrie A (1978) Individuality in Pain and Suffering. Chicago: University of Chicago Press.
- Avegno EM, Lobell TD, Itoga CA, Baynes BB, Whitaker AM et al. (2018) Central Amygdala Circuits Mediate Hyperalgesia in Alcohol-Dependent Rats. J Neurosci 38: 7761-7773. [Crossref]
- Brown RA, Cutter HS (1977) Alcohol, customary drinking behavior, and pain. J Abnorm Psychol 86: 179-188. [Crossref]
- Stewart SH, Finn PR, Pihl RO (1995) A dose-response study of the effects of alcohol on the perceptions of pain and discomfort due to electric shock in men at high familial-genetic risk for alcoholism. Psychopharmacology (Berl) 119: 261-267. [Crossref]
- Egli M, Koob GF, Edwards S (2012) Alcohol dependence as a chronic pain disorder. Neurosci Biobehav Rev 36: 2179-2192. [Crossref]
- Kimpel MW, Brown MM, Froehlich JC (2003) Pain thresholds in alcohol preferring and non-preferring rats: diurnal and repeated trial line differences. Alcohol Clin Exp Res 27: 1921-1928. [Crossref]
- Chester JA, Price CS, Froehlich JC (2002) Inverse genetic association between alcohol preference and severity of alcohol withdrawal in two sets of rat lines selected for the same phenotype. Alcohol Clin Exp Res 26: 19-27. [Crossref]
- Kampov Polevoy AB (1996) Pain sensitivity and saccharin intake in alcohol-preferring and -nonpreferring rat strains. Physiol Behav 59: 683-688.
- Mogil JS, Wilson SG, Bon K, Lee SE, Chung K et al. (1999) Heritability of nociception II. 'Types' of nociception revealed by genetic correlation analysis. Pain 80: 83-93. [Crossref]
- Shir Y, Seltzer Z (1990) A-fibers mediate mechanical hyperesthesia and allodynia and C-fibers mediate thermal hyperalgesia in a new model of causalgiform pain disorders in rats. Neurosci Lett 115: 62-67. [Crossref]
- Lima D, Avelino A (1994) Spinal c-fos expression is differentially induced by brief or persistent noxious stimulation. Neuroreport 5: 1853-1856. [Crossref]
- Lenz FA, Gracely RH, Rowland LH, Dougherty PM (1994) A population of cells in the human thalamic principal sensory nucleus respond to painful mechanical stimuli. Neurosci Lett 180: 46-50. [Crossref]
- Hwang BH, Stewart R, Zhang JK, Lumeng L, Li TK (2004) Corticotropin-releasing factor gene expression is down-regulated in the central nucleus of the amygdala of alcohol-preferring rats which exhibit high anxiety: a comparison between rat lines selectively bred for high and low alcohol preference. Brain Res 1026: 143-150. [Crossref]
- Zhang H, Sakharkar AJ, Shi G, Ugale R, Prakash A et al. (2010) Neuropeptide Y Signaling in the Central Nucleus of Amygdala Regulates Alcohol-Drinking and Anxiety-Like Behaviors of Alcohol-Preferring Rats. Alcohol Clin Exp Res 34: 451-461. [Crossref]
- Pettinati HM (2004) Antidepressant treatment of co-occurring depression and alcohol dependence. Biol Psychiatry 56: 785-792. [Crossref]
- Cornelius JR, Maisto SA, Martin CS, Bukstein OG, Salloum IM et al. (2004) Major depression associated with earlier alcohol relapse in treated teens with AUD. Addict Behav 29: 1035-1038. [Crossref]
- Korpi ER, Sinclair JD, Kaheinen P, Viitamaa T, Hellevuo K et al. (1988) Brain regional and adrenal monoamine concentrations and behavioral responses to stress in alcohol-preferring AA and alcohol-avoiding ANA rats. Alcohol 5: 417-425. [Crossref]
- Ehlers CL, Walker BM, Pian JP, Roth JL, Slawecki CJ (2007) Increased alcohol drinking in isolate-housed alcohol-preferring rats. Behav Neurosci 121: 111-119. [Crossref]