Overwhelming Evidence Transplacental Transmission of Human Papillomavirus Primarily Causes Autism
A B S T R A C T
Because the concordance rate between identical twins is only 88%, an environmental factor must cause autism spectrum disorder (ASD). Furthermore, when identical twins share ASD, it is to varying degrees suggesting different prenatal environments exist, which occurs when identical twins have separate placentas (~30% of the time). Placental inclusions are predictive of ASD along with excessive increases in extra-axial cerebral spinal fluid (CSF) detected by MRI in the brains of 6- and 12-month-old infants later diagnosed at 2 years with ASD. The human papillomavirus (HPV) can infect the trophoblast cells of placentas and transmit to the fetus where it infects the epithelial cells of the choroid plexus, a centrally located lining inside the brain responsible for producing CSF via the SLC4A10 gene product. HPV causes epigenetic changes, deletions, and duplications of genes, and besides its characteristic methylation patterns, the SLC4A10 gene was found to be increased in children with ASD. Moreover, male placentas implant close to the cervix (low-lying) three times more often than female placentas paralleling the ASD ratio of ~3:1 (boys to girls). Finally, the Australian HPV vaccination programme that began in 2007 might explain why the 0-4 yr. ASD incidence did not increase from 2010 to 2015.
Keywords
Autism spectrum disorder, cerebral spinal fluid, choroid plexus, human papillomavirus, placenta, prenatal environment
Introduction
Autism spectrum disorder (ASD) encompasses a wide range of neurological developmental conditions usually characterized by insufficiencies in social interactions and communication abilities. Everything from genetics (including epigenetic changes, copy number variants, somatic mutations, and polymorphisms) to environmental exposures (including low vitamin D levels, vaccinations, chemicals, medications, drugs, and cytokines) and combinations of both have been frantically investigated in a desperate attempt to stop the alarming rate increase. The alarming rate increase was blamed on enhanced public awareness and changes in diagnostic criteria, but the rate of increase in the ASD incidences around the world over the past few decades were shown to be real by the CHARGE study [1, 2]. The rate varies in developed countries around the world from 1 in 27 in Hong Kong to 1 in 3,333 in Poland, and in the United States (U.S.) it may have been as high as 1 in 45 in 2017 [3, 4]. ASD is pandemic and thought to be genetic in origin, but because it increased at exponential rates in all races around the world in recent decades suggests it is primarily due to an environmental factor like a contagious disease.
I Environment Versus Genetics
The best evidence an environmental factor causes ASD is provided by identical twin studies. Identical twins share identical genetics, but they have less than a 100% concordance rate (both have it) for ASD. We expect pairwise concordance rates of 100% for identical twins if ASD is a genetic disorder, but only an 88% pairwise concordance rate exists between identical twins and a 31% pairwise concordance rate exists between non-identical or fraternal twins as shown by a large twin study involving 277 twin pairs [5]. Another study of 366 identical twin pairs demonstrated the proband concordance rate for ASD was as high as ~96%, but the similarity of shared symptoms was low; the authors concluded some non-shared environmental factor during early development caused the broad range of genetic and behavioural differences observed between identical twins [6]. If the etiology of ASD was genetic, the concordance rate between identical twins would be 100% and their symptoms would also be 100% identical, but they are not showing ASD is caused by an environmental factor. Twin data including non-identical twins with concordance rates ~31% shows blood-borne substances like alcohol, medications, cytokines, infections, or low prenatal vitamin D levels cannot be the primary cause of ASD because all twins - identical or not - would share that exposure equally and would have a 100% concordance rate and the same degree of symptoms, but this is not the case [7].
Blood-borne viruses like HIV, Herpes simplex virus, cytomegalovirus, and Epstein Barr Virus can infect the mother but cannot be the primary cause of ASD because both non-identical and identical twins would have a 100% concordance rate for those viruses [8-10]. Some infections in utero are associated with an increased risk for ASD and would cause both twins to be concordant, but these types of infections do not explain the discordant rates of ASD between identical twins [11]. The primary cause of ASD must explain why the concordance rate of ASD is less than 100% for identical twins who have identical DNA. The prenatal environment of identical twins is only the same 70% of the time when they share the same placenta (monochorionic); identical twin prenatal environments are different 30% of the time when they have separate placentas and non-identical twins’ prenatal environment is always different because they always have separate placentas (dichorionic) [12].
When the placenta implants close to a contaminated cervix, it can become infected with HPV and transmit it to the fetus [13, 14]. Placentas can implant near the cervix (low-lying), but they usually move upward in the uterus during the second or third trimester. Of the low-lying placentas, about 90% clear the cervix area by 32 weeks and about 96% clear the cervix by 36 weeks circumventing placenta previa [15, 16]. Identical twins will be concordant for ASD only if either their shared placenta (70%) is located near the infected cervix and both twins become infected to the same degree with similar symptoms (top of Figure 1) or both non-shared placentas (30%) are near the infected cervix but one is closer to it than the other and becomes more infected than the other with increased ASD symptoms (bottom of Figure 1). The identical twins will be discordant for ASD if only one placenta is near the infected cervix while the other is not (middle of Figure 1). For non-identical twins to be concordant for ASD, both placentas must be close to the infected cervix (top of Figure 2), because if only one placenta is close to the infected cervix, they will be discordant for ASD (bottom of Figure 2). The probability of both identical twin placentas locating close to the cervix is estimated from the non-identical twin concordance rate of 31% [5]. The total concordance rate can be estimated for identical twins by multiplying 31% (probability of both placentas locating near the cervix) by 30% (non-shared placentas) which gives about 10% that is added to 70% (shared placentas locating near the cervix) for an estimated 80% pairwise concordance rate. That is within 10% of the observed 88% concordance rate for identical twins [5] which is acceptable because some smaller studies found lower concordance rates for identical twins. And the fact that males tend to have low-lying placentas three times more often than females, demonstrated by their placenta previa rates, matches the averaged worldwide ratio of boys to girls with ASD (~3:1) [4, 17, 18].
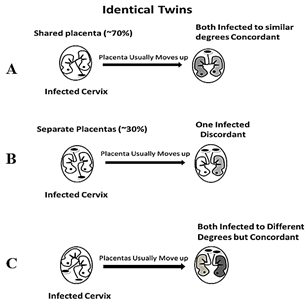
Figure 1: Identical twins have three scenarios for placentas: A) 70% shared placentas – concordant, B) 30% non-shared or separate placentas and only one is low in the uterus – discordant, and C) both separate placentas are low in the uterus and can be infected to different degrees – concordant.
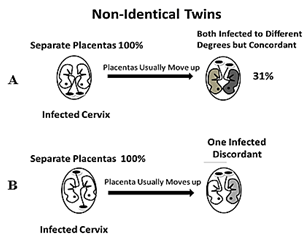
Figure 2: Non-identical twins never share a placenta so they only have two scenarios: A) Both placentas are low in the uterus and can be infected to different degrees – concordant (31%), and B) only one placenta is low in the uterus – discordant.
The most prevalent pandemic cervical infection is the human papillomavirus (HPV) [19]. The exponential increase in the incidence of HPV+ oropharyngeal cancers in the U.S. (Figure 3A) parallels the exponential increase in ASD in the U.S. (Figure 3B) during the same time frame, and in Europe (Sweden) the HPV+ oropharyngeal cancers (Figure 4A) also parallel the exponential increase in ASD (Figure 4B) [20, 21]. Clinicians find HPV in the trophoblast cells of placentas that can transmit HPV to the fetus [13, 14, 22, 23]. Trophoblast inclusions are predictive of ASD and placental abnormalities and preterm births are major risk factors for ASD [24, 25]. Unfortunately, clinicians have not yet tested for HPV in the trophoblast cells with inclusions, but HPV is known to cause placental abnormalities, preterm births, low birth weights, and spontaneous abortions [26, 27]. The severity of ASD symptoms might depend on the viral load due to the location of the placenta, i.e., how close the placenta embeds near the contaminated cervix, the timing of infection (first trimester or longer), the strain(s) of HPV (e.g., -6, -16, -52, -90, etc.), or some or all of those factors combined which might explain why a broad spectrum of autistic symptoms exists.
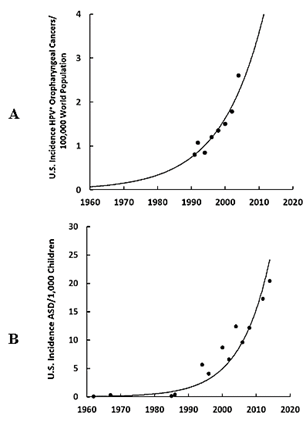
Figure 3: A) Temporal incidence in the U.S. of HPV+ oropharyngeal cancers per 100,000 world population, B) Temporal incidence in the U.S. of ASD per 1,000 children. Note that if more than one study existed, the ASD incidences were averaged [21].
II Worldwide Prevalence of Cervical HPV Infection, ASD, and Placenta Previa
The prevalence of HPV infection varies by the mother’s age, continent, and region [28]. Older women have a much lower HPV prevalence than younger women of child-bearing age. The continent of Africa (21.1%) had the highest HPV prevalence followed by Europe (14.2%), and North and South America (11.5%; including the regions of Canada, U.S., Greenland, Central and South Americas, and the Caribbean islands), and Asia (9.4%); while Australia was excluded because they have an ongoing HPV vaccination programme. The order of HPV prevalence is Africa (21.1%)> Europe (14.2%) > Americas (11.5%) [N. America (4.7%)] >Asia (9.4%).
The order of HPV prevalence does not parallel the order of ASD prevalence found around the world [3]. The ASD rate in Africa was not determined, but in Europe the rate was about 0.681%. The South American, Central American, and Greenland ASD rates were not determined, but they were determined in the U.S. as 1 in 45 (2.22%) and in Canada as 1 in 94 (1.06%). The ASD rate in North America cannot be calculated because the rate was not determined in Greenland but if it is assumed to be similar to Europe’s rate (0.681%), North America would be about 1.32%. The ASD rates were only determined in eastern Asia at 1 in 38 (2.63%) in South Korea, 1 in 55 (1.82%) in Japan, and 1 in 435 (0.23%) in China for an average of 1.56%. The order for ASD prevalence is Asia (~1.56%) > N. America (~1.32%) > Europe (~0.681%, excluding southern regions) which is the opposite order of HPV prevalence in similar regions of the world.
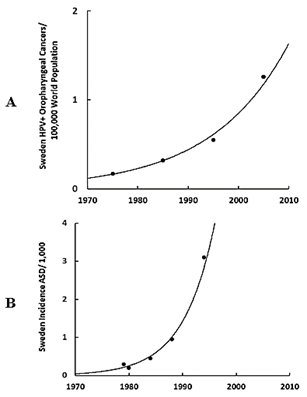
Figure 4: A) Temporal incidence in Europe (Sweden) of HPV+ oropharyngeal cancers per 100,000 world population, B) Temporal incidence in Europe (Sweden) of ASD per 1,000 children.
But cervical HPV infection does not predict the rate of ASD, because the placenta cannot get infected unless it is located near the cervix. Thus, placenta previa rates would be the best predictors of ASD because they reflect the rate of placentas implanting low in the uterus where they can become infected by HPV. Up to 30% of placentas can be low-lying in the uterus prior to 6 months gestation but sometime before delivery 90% move higher up in the uterus avoiding placenta previa [29, 30]. The placenta previa rates were found to be the highest in Asia (12.2 per 1000 pregnancies) followed by Europe (3.6 per 1000 pregnancies) and North America (2.9 per 1000 pregnancies) with the lowest rates in Sub-Saharan Africa (2.7 per 1000 pregnancies) [31]. The region of the world can reflect contributions towards placenta previa rates from genetics (race/ethnicity) or something in the environment (e.g., water, food, or chemical exposures). A study in northern California involving different races living in the same location shows genetics plays the largest role in placenta previa rates [32]. The placenta previa rates correlate well with the ASD rates as the Asians have the highest placenta previa rates of all the races and also have the highest ASD rates in the world, e.g., Hong Kong (1 in 27 or 3.7%), South Korea (1 in 38 or 2.63%), and Japan (1 in 55 or 1.82%). The placenta previa rates parallel the ASD rates: Asians (0.64%) > Native Americans (0.6%) > African Americans (0.44%) > Caucasians (0.36%) > Hispanics (0.34%).
III Can HPV Infect the Brain?
HPV was found in neurons and in the post-mortem brains of children who had focal cortical dysplasia type IIB (epilepsy and seizures), which is also sometimes associated with ASD [33-36]. Of the five HPV genera (alpha, beta, gamma, mu, and nu), two genera (beta and gamma) can infect epithelial cells [37]. The choroid plexus, which is a centrally located lining inside the brain that produces cerebral spinal fluid (CSF), is comprised of epithelial cells that have the sodium-driven chloride bicarbonate exchanger, SLC4A10 gene product, required for CSF production [38, 39]. CSF is secreted when SCL4A10 unidirectionally transports sodium, chlorine, and bicarbonate from the blood to the ventricles of the brain. SLC4A10 disruption enhances neuronal excitability, modifies synaptic short-term plasticity, and is sometimes associated with complex partial epilepsy and mental retardation [40, 41]. Most importantly, increases in extra-axial CSF at 6 and 12 months of age observed in brain ventricles by MRI were predictive of infants who developed ASD at 2 yrs. of age, and increasing amounts of CSF correlated with increasing severity of ASD symptoms [42]. An increased copy number of the SLC4A10 gene is associated with ASD and integration of HPV into the genome results in duplications and deletions of genes [43, 44]. More scientific evidence that HPV causes problems in the brain is obtained from the papillomas and tumors of the choroid plexus found in children younger than 2 [45, 46]. The E6 and E7 oncogene proteins of HPV caused tumors in the choroid plexus of transgenic mice [38]. Further, neuroinflammation is a hallmark of ASD and can be caused by activation of the neuroglial cells that remove infections in the brain [47, 48]. Moreover, the gut is similar to the choroid plexus, having similar immune functions and a single layer of epithelial cells that HPV can infect [49]. In fact, 75% of ASD children have gastrointestinal diseases like inflammatory bowel disease [50].
IV Evidence of Biochemical and Epigenetic Fingerprints of HPV in ASD Children
Because HPV is involved in cancer, we would expect to find biochemical similarities between ASD and cancer, and indeed they exist (see Table 1 for the following discussion) [51-53]. Both ASD and cancer have disruptions in the PI3K-Akt-mTOR signaling pathway; mTOR serine/threonine kinase mediates signaling pathways required for glial and neuronal differentiation in brain development. mTORC1 signaling is disrupted in ASD and occurs in pediatric brain tumors, seizures, learning disabilities, and mental retardation [54]. mTOR signaling is activated through PTEN in children with ASD and the E6 protein of HPV activates mTORC1 signaling [55, 56]. Clinicians use Rapamycin, which inhibits the mTORC1 pathway, to treat both ASD and cancer [57-59]. More biochemical evidence is obtained from the UBE3A gene that encodes the ubiquitin E3 ligase, E6-A, which degrades p53 (the Guardian of the Genome) and is associated with both ASD and HPV [60, 61].
Table
1:
Endpoints in common between HPV and ASD.
|
Endpoint |
HPV |
ASD |
|
Placental Inclusions |
HPV not tested but viral infections are
known to cause inclusions [85] |
Predictive of ASD [24] |
|
mTOR |
E6 protein activates mTORC1 [56] (Rapamycin used medically to inhibit [59]) |
mTOR activated [55] (Rapamycin used medically to inhibit [57,
58]) |
|
UBE3A |
Duplicates gene that degrades p53
[61] causing genomic instability in cervical cancer [74] |
Gene is duplicated in ASD [60, 62] causing
genomic instability [75] and cancers |
|
SLC4A10 |
Duplicates gene [44] |
Duplicated gene [62] increases CSF in brain
recently related to ASD [42] |
|
Cancer |
Causes several types of cancers (including
children’s choroid plexus brain tumors [46] and papillomas [45]) |
Increased incidence of cancers in the ASD
population [63, 64] |
|
Preeclampsia |
Infection increases blood pressure of
pregnant women [66] |
A known risk factor for ASD [65] |
|
DNA Methylation/ genomic instability |
Abnormal in cervical and only HPV+
oropharyngeal cancers [71, 72]/ genomic instability [74] |
Identical twins discordant for DNA
methylation [70]/genomic instability [75] |
UBE3A is another consistently duplicated gene in ASD besides SLC4A10 [62]. Further evidence HPV probably causes ASD is obtained from significantly higher incidences of cancer in the ASD population [63, 64]. Moreover, preeclampsia (high blood pressure) in associated with pregnant women who had an ASD child and an almost 2-fold increase in the risk for developing preeclampsia exists for mothers with HPV infection [65, 66]. Finally, ASD children all had lower vitamin D3 levels, which were only lower in the discordant identical twin who had ASD even though both twins had similar diets and sun exposures [67]. HPV may decrease vitamin D levels by increasing the production of the enzyme alpha-N-acetylgalactosaminidase that was found to be elevated in ASD children [68]. Alpha-N-acetylgalactosaminidase cleaves the sugar moiety from the vitamin D binding protein so that it can no longer bind to and transport vitamin D into the bloodstream. Another detrimental effect of cleaving off the sugar moiety is losing enzymatic conversion to the macrophage-activating factor, which results in suppression of the innate immune response. Vitamin D is also required for T-cell activation and antigen receptor signaling, so that low levels suppress the adaptive immune response as well [69]. Increasing that enzyme, results in suppression of both the innate and adaptive immune responses allowing HPV to escape immune surveillance.
Epigenetic events like DNA methylation of promoters, duplications and deletions of genes, and point mutations can make ASD appear to be genetic disguising the environmental factor involved. Decreases or increases in the production of gene products occurs when promotors are either hypomethylated or hypermethylated. An abnormal DNA methylation pattern is only found in the identical twin with ASD - not in the discordant twin– and an abnormal DNA methylation pattern is also found in cervical cancer and only in HPV positive not negative oropharyngeal cancers [70-72]. Over 190 sites in the host genome can allow HPV integration that results in duplications and deletions of genes resulting in genomic instability observed in cervical cancers and in ASD [73-75]. Some point mutations found in ASD might be from deamination of methylated cytosines in CpHpG sites (where H = A, C or T) [76].
Another potential genetic indicator was thought to be the higher occurrence of ASD between siblings, but over half the infected women could not clear HPV for 2 years so that another pregnancy during that time could also yield a child with ASD [77]. One year between pregnancies was found to be three times riskier than 3 years between pregnancies for having a second child with ASD [78]. If ASD had a genetic origin, the time between sibling births would not matter. In addition to short spacing times between sibling births (<2 yrs.), long spacing times (>6 yrs.) also increased the risk for having a child with ASD; however, that could be due either to reinfection or to the mother becoming immunosuppressed and HPV becoming reactivated [79, 80]. Vitamin D3 deficiency (and insufficiency) is one way to cause immunosuppression and reactivate HPV [81]. Additionally, gestational vitamin D deficiency and neonatal or combined maternal and neonatal insufficient levels of vitamin D also represent an increased risk for ASD [7, 82].
V Prevention
The best strategy to prevent ASD is for people, especially women, to get HPV vaccinated (3 shots) prior to and even during pregnancy, which was found to be safe, because the mother can supply passive immunity to the neonate and infant [83]. Australia began a free school HPV vaccination programme (quadrivalent HPV-6, -11, -16, -18) for 12-13 yr. girls in 2007 that was extended to boys in 2013. Vaccinated 12–13-year-old girls in 2007 were of child-bearing age in 2015 (20-21 yr.) and some had children 4 yrs. old or younger and an Australian study found no increase in the incidence of ASD from 2010 to 2015 for the 0-4 yr. age group [18]. Besides HPV vaccination, people and children should also increase their vitamin D3 to sufficient blood levels (≥50 nmol/L) to boost their innate and adaptive immune responses and improve their brain health [84].
Conflicts of Interest
None.
Acknowledgement
None.
Funding
None.
Disclosure
The findings and conclusions in this paper have not been formally disseminated by the Food and Drug Administration and should not be construed to represent any agency determination or policy. The mention of commercial products, their sources, or their use in connection with material reported herein is not to be construed as either an actual or implied endorsement of such products by Department of Health and Human Services.
Abbreviation
ASD: Autism Spectrum Disorder
CSF: Cerebral Spinal Fluid
HPV: Human Papillomavirus
U.S.: United States
Article Info
Article Type
Review ArticlePublication history
Received: Fri 26, Nov 2021Accepted: Sat 11, Dec 2021
Published: Fri 31, Dec 2021
Copyright
© 2023 Dianne E. Godar. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.NNB.2021.04.02
Figures & Tables
Table
1:
Endpoints in common between HPV and ASD.
|
Endpoint |
HPV |
ASD |
|
Placental Inclusions |
HPV not tested but viral infections are
known to cause inclusions [85] |
Predictive of ASD [24] |
|
mTOR |
E6 protein activates mTORC1 [56] (Rapamycin used medically to inhibit [59]) |
mTOR activated [55] (Rapamycin used medically to inhibit [57,
58]) |
|
UBE3A |
Duplicates gene that degrades p53
[61] causing genomic instability in cervical cancer [74] |
Gene is duplicated in ASD [60, 62] causing
genomic instability [75] and cancers |
|
SLC4A10 |
Duplicates gene [44] |
Duplicated gene [62] increases CSF in brain
recently related to ASD [42] |
|
Cancer |
Causes several types of cancers (including
children’s choroid plexus brain tumors [46] and papillomas [45]) |
Increased incidence of cancers in the ASD
population [63, 64] |
|
Preeclampsia |
Infection increases blood pressure of
pregnant women [66] |
A known risk factor for ASD [65] |
|
DNA Methylation/ genomic instability |
Abnormal in cervical and only HPV+
oropharyngeal cancers [71, 72]/ genomic instability [74] |
Identical twins discordant for DNA
methylation [70]/genomic instability [75] |
References
1. Hansen SN, Schendel
DE, Parner ET (2015) Explaining the increase in the prevalence of autism
spectrum disorders: the proportion attributable to changes in reporting
practices. JAMA Pediatr 169: 56-62. [Crossref]
2. Hertz-Picciotto I,
Croen LA, Hansen R, Jones CR, van de Water J et al. (2006) The CHARGE study: an
epidemiologic investigation of genetic and environmental factors contributing
to autism. Environ Health Perspect 114: 1119-1125. [Crossref]
3. https://www.focusforhealth.org/autism-rates-across-the-developed-world/
Assessed on: 24/07/2018
4. Xu G, Strathearn L,
Liu B, Bao W (2018) Prevalence of Autism Spectrum Disorder Among US Children
and Adolescents, 2014-2016. JAMA 319: 81-82. [Crossref]
5. Rosenberg RE, Law
JK, Yenokyan G, McGrady J, Kaufmann WE et al. (2009) Characteristics and concordance
of autism spectrum disorders among 277 twin pairs. Arch Pediatr Adolesc Med
163: 907-914. [Crossref]
6. Castelbaum L,
Sylvester CM, Zhang Y, Yu Q, Constantino JN (2020) On the Nature of Monozygotic
Twin Concordance and Discordance for Autistic Trait Severity: A Quantitative
Analysis. Behav Genet 50: 263-272. [Crossref]
7. Vinkhuyzen AAE,
Eyles DW, Burne THJ, Blanken LME, Kruithof CJ et al. (2018) Gestational vitamin
D deficiency and autism-related traits: the Generation R Study. Mol
Psychiatry 23: 240-246. [Crossref]
8. Musetti L, Albizzati A, Grioni A, Rossetti M, Saccani M et al. (1993) Autistic
disorder associated with congenital HIV infection. Eur Child Adolesc
Psychiatry 2: 221-225. [Crossref]
9. Mahic M, Mjaaland
S, Bøvelstad HM, Gunnes N, Susser E et al. (2017) Maternal Immunoreactivity to
Herpes Simplex Virus 2 and Risk of Autism Spectrum Disorder in Male Offspring. mSphere
2: e00016- e00017. [Crossref]
10. Valayi S,
Eftekharian MM, Taheri M, Alikhani MY (2017) Evaluation of antibodies to
cytomegalovirus and Epstein-Barr virus in patients with autism spectrum
disorder. Hum Antibodies 26: 165-169. [Crossref]
11. Al-Haddad BJS,
Jacobsson B, Chabra S, Modzelewska D, Olson EM et al. (2019) Long-term Risk of
Neuropsychiatric Disease After Exposure to Infection In Utero. JAMA
Psychiatry 76: 594-602. [Crossref]
12. Faye-Petersen OM,
Heller DS, Joshi VV (2006) Handbook of Placental Pathology, 2nd edition.
London: Taylor & Francis.
13. Rombaldi RL,
Serafini EP, Mandelli J, Zimmermann E, Losquiavo KP (2008) Transplacental
transmission of human papillomavirus. Virol J 5: 106. [Crossref]
14. Smith EM, Parker
MA, Rubenstein LM, Haugen TH, Hamsikova E et al. (2010) Evidence for vertical
transmission of HPV from mothers to infants. Infect Dis Obstet Gynecol
2010: 326369. [Crossref]
15. Heller HT, Mullen
KM, Gordon RW, Reiss RE, Benson CB (2014) Outcomes of pregnancies with a
low-lying placenta diagnosed on second-trimester sonography. J Ultrasound
Med 33: 691-696. [Crossref]
16. Wen SW, Demissie K,
Liu S, Marcoux S, Kramer MS (2000) Placenta praevia and male sex at birth:
results from a population-based study. Paediatr Perinat Epidemiol 14:
300-304. [Crossref]
17. Köstü B, Ercan Ö,
Özer A, Bakacak M, Avcı F (2015) Male fetus domination in total placenta previa
cases. Perinat J 23: 84-88.
18. May T, Williams K
(2018) Brief Report: Gender and Age of Diagnosis Time Trends in Children with
Autism Using Australian Medicare Data. J Autism Dev Disord 48:
4056-4062. [Crossref]
19. Human
papillomavirus (HPV) and cervical cancer (2020) World Health Organization.
20. Chaturvedi AK,
Engels EA, Pfeiffer RM, Hernandez BY, Xiao W et al. (2011) Human papillomavirus
and rising oropharyngeal cancer incidence in the United States. J Clin Oncol
29: 4294-4301. [Crossref]
21. Autism Spectrum
Disorder (ASD), Summary of Autism Spectrum Disorder (ASD) Prevalence Studies.
Centers for Disease Control and Prevention.
22. You H, Liu Y,
Agrawal N, Prasad CK, Edwards JL et al. (2008) Multiple human papillomavirus
types replicate in 3A trophoblasts. Placenta 29: 30-38. [Crossref]
23. Weyn C, Thomas D,
Jani J, Guizani M, Donner C et al. (2011) Evidence of human papillomavirus in
the placenta. J Infect Dis 203: 341-343. [Crossref]
24. Walker CK, Anderson
KW, Milano KM, Ye S, Tancredi DJ et al. (2013) Trophoblast inclusions are
significantly increased in the placentas of children in families at risk for
autism. Biol Psychiatry 74: 204-211. [Crossref]
25. Placental function
linked to brain injuries associated with autism (2019) Children's National
Health System. ScienceDaily.
26. Zuo Z, Goel S,
Carter JE (2011) Association of cervical cytology and HPV DNA status during
pregnancy with placental abnormalities and preterm birth. Am J Clin Pathol
136: 260-265. [Crossref]
27. Fezer GF, de Matos
MB, Nau AL, Zeigelboim BS, Marques JM et al. (2017) PERINATAL FEATURES OF
CHILDREN WITH AUTISM SPECTRUM DISORDER. Rev Paul Pediatr 35: 130-135. [Crossref]
28. Bruni L, Diaz M, Barrionuevo-Rosas L, Herrero R, Bray F et al. (2016) Global
estimates of human papillomavirus vaccination coverage by region and income
level: a pooled analysis. Lancet Glob Health 4: e453- e463. [Crossref]
29. Khan AT, Stewart KS
(1987) Ultrasound placental localisation in early pregnancy. Scott Med J
32: 19-21. [Crossref]
30. Rizos N, Doran TA,
Miskin M, Benzie RJ, Ford JA (1979) Natural history of placenta previa
ascertained by diagnostic ultrasound. Am J Obstet Gynecol 133: 287-291.
[Crossref]
31. Cresswell JA,
Ronsmans C, Calvert C, Filippi V (2013) Prevalence of placenta praevia by world
region: a systematic review and meta-analysis. Trop Med Int Health 18:
712-724. [Crossref]
32. Kim LH, Caughey AB,
Laguardia JC, Escobar GJ (2012) Racial and ethnic differences in the prevalence
of placenta previa. J Perinatol 32: 260-264. [Crossref]
33. Füle T, Máthé M,
Suba Z, Csapó Z, Szarvas T et al. (2006) The presence of human papillomavirus
16 in neural structures and vascular endothelial cells. Virology 348:
289-296. [Crossref]
34. Chen J, Tsai V,
Parker WE, Aronica E, Baybis M et al. (2012) Detection of human papillomavirus
in human focal cortical dysplasia type IIB. Ann Neurol 72: 881-892. [Crossref]
35. Millichap JG (2013)
Focal Cortical Dysplasia Type IIB and Human Papillomavirus. Pediatr Neurol
Briefs 27: 24-24.
36. Casanova MF, El-Baz
AS, Kamat SS, Dombroski BA, Khalifa F et al. (2013) Focal cortical dysplasias
in autism spectrum disorders. Acta Neuropathol Commun 1: 67. [Crossref]
37. de Villiers EM,
Fauquet C, Broker TR, Bernard HU, zur Hausen H (2004) Classification of
papillomaviruses. Virology 324: 17-27. [Crossref]
38. Arbeit JM, Münger
K, Howley PM, Hanahan D (1993) Neuroepithelial carcinomas in mice transgenic
with human papillomavirus type 16 E6/E7 ORFs. Am J Pathol 142:
1187-1197. [Crossref]
39. Praetorius J,
Nejsum LN, Nielsen S (2004) A SCL4A10 gene product maps selectively to the
basolateral plasma membrane of choroid plexus epithelial cells. Am J Physiol
Cell Physiol 286: C601-610. [Crossref]
40. Sinning A, Liebmann
L, Hübner CA (2015) Disruption of Slc4a10 augments neuronal excitability and
modulates synaptic short-term plasticity. Front Cell Neurosci 9: 223. [Crossref]
41. Gurnett CA, Veile
R, Zempel J, Blackburn L, Lovett M (2008) Disruption of sodium bicarbonate
transporter SLC4A10 in a patient with complex partial epilepsy and mental
retardation. Arch Neurol 65: 550-552. [Crossref]
42. Shen MD, Kim SH,
McKinstry RC, Gu H, Hazlett HC et al. (2017) Increased Extra-axial
Cerebrospinal Fluid in High-Risk Infants Who Later Develop Autism. Biol
Psychiatry 82: 186-193. [Crossref]
43. Schmitz M, Driesch
C, Jansen L, Runnebaum IB, Dürst M (2012) Non-random integration of the HPV
genome in cervical cancer. PLoS One 7: e39632. [Crossref]
44. Sebat J, Lakshmi B,
Malhotra D, Troge J, Lese-Martin C et al. (2007) Strong association of de novo
copy number mutations with autism. Science 316: 445-449. [Crossref]
45. Prasad GL,
Mahapatra AK (2015) Case series of choroid plexus papilloma in children at
uncommon locations and review of the literature. Surg Neurol Int 6: 151.
[Crossref]
46. Ogiwara H, Dipatrim
AJ Jr, Aldenm TD, Bowmanm RM, Tomita T (2012) Choroid plexus tumors in
pediatric patients. Br J Neurosurg 26: 32-37. [Crossref]
47. Vargas DL,
Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA (2005) Neuroglial activation
and neuroinflammation in the brain of patients with autism. Ann Neurol
57: 67-81. [Crossref]
48. Shastri A, Bonifati
DM, Kishore U (2013) Innate immunity and neuroinflammation. Mediators Inflamm
2013: 342931. [Crossref]
49. de Jonge WJ (2013)
The Gut's Little Brain in Control of Intestinal Immunity. ISRN Gastroenterol
2013: 630159. [Crossref]
50. Horvath K, Perman
JA (2002) Autistic disorder and gastrointestinal disease. Curr Opin Pediatr
14: 583-587. [Crossref]
51. Godar DE, Merrill
SJ (2017) Untangling the most probable role for vitamin D 3 in autism. Dermatoendocrinol
9: e1387702. [Crossref]
52. Godar DE, Gurov R,
Merrill SJ (2017) All sites but skin cancer incidences analyzed worldwide by
sex, age, and skin type over time (1955-2007), advancing age, and UVB dose
reveals important carcinogenic drivers J Epi Res 3: 65-80.
53. Crawley JN, Heyer
WD, LaSalle JM (2016) Autism and Cancer Share Risk Genes, Pathways, and Drug
Targets. Trends Genet 32: 139-146. [Crossref]
54. Lee DY (2015) Roles
of mTOR Signaling in Brain Development. Exp Neurobiol 24: 177-185. [Crossref]
55. Tilot AK, Frazier
2nd TW, Eng C (2015) Balancing Proliferation and Connectivity in
PTEN-associated Autism Spectrum Disorder. Neurotherapeutics 12: 609-619.
[Crossref]
56. Spangle JM, Munger
K (2010) The human papillomavirus type 16 E6 oncoprotein activates mTORC1
signaling and increases protein synthesis. J Virol 84: 9398-9407. [Crossref]
57. Crino PB (2016) The
mTOR signalling cascade: paving new roads to cure neurological disease. Nat
Rev Neurol 12: 379-392. [Crossref]
58. Sato A (2016) mTOR,
a Potential Target to Treat Autism Spectrum Disorder. CNS Neurol Disord Drug
Targets 15: 523-543. [Crossref]
59. Coppock JD, Wieking
BG, Molinolo AA, Gutkind JS, Miskimins KW et al. (2013) Improved clearance
during treatment of HPV-positive head and neck cancer through mTOR inhibition. Neoplasia
15: 620-630. [Crossref]
60. Yi JJ, Berrios J,
Newbern JM, Snider WD, Philpot BD et al. (2015) An Autism-Linked Mutation
Disables Phosphorylation Control of UBE3A. Cell 162: 795-807. [Crossref]
61. Brimer N, Lyons C,
Pol SBV (2007) Association of E6AP (UBE3A) with human papillomavirus type 11 E6
protein. Virology 358: 303-310. [Crossref]
62. Bourgeron T (2015)
From the genetic architecture to synaptic plasticity in autism spectrum
disorder. Nat Rev Neurosci 16: 551-563. [Crossref]
63. Crespi B (2011)
Autism and cancer risk. Autism Res 4: 302-310. [Crossref]
64. Chiang HL, Liu CJ,
Hu YW, Chen SC, Hu LY et al. (2015) Risk of cancer in children, adolescents,
and young adults with autistic disorder. J Pediatr 166: 418-423. [Crossref]
65. Walker CK,
Krakowiak P, Baker A, Hansen RL, Ozonoff S et al. (2015) Preeclampsia,
placental insufficiency, and autism spectrum disorder or developmental delay. JAMA
Pediatr 169: 154-162. [Crossref]
66. McDonnold M, Dunn
H, Hester A, Pacheco LD, Hankins et al. (2014) High risk human papillomavirus
at entry to prenatal care and risk of preeclampsia. Am J Obstet Gynecol
210: 138.e1-138.e5. [Crossref]
67. Fahmy F, Sabri NA,
El Hamamsy MH, El Sawi M, Zaki OK (2016) VITAMIN D INTAKE AND SUN EXPOSURE IN AUTISTIC
CHILDREN. Int J Pharm Sci Res 7: 1043-1049.
68. Reddi AL,
Sankaranarayanan K, Arulraj HS, Devaraj N, Devaraj H (2000) Serum alpha-N-
acetylgalactosaminidase is associated with diagnosis/prognosis of patients with
squamous cell carcinoma of the uterine cervix. Cancer Lett 158: 61-64. [Crossref]
69. von Essen MR,
Kongsbak M, Schjerling P, Olgaard K, Odum N et al. (2010) Vitamin D controls T
cell antigen receptor signaling and activation of human T cells. Nat Immunol
11: 344-349. [Crossref]
70. Wong CCY, Meaburn
EL, Ronald A, Price TS, Jeffries AR et al. (2014) Methylomic analysis of
identical twins discordant for autism spectrum disorder and related behavioural
traits. Mol Psychiatry 19: 495-503. [Crossref]
71. Siegel EM, Riggs
BM, Delmas AL, Koch A, Hakam A et al. (2015) Quantitative DNA methylation
analysis of candidate genes in cervical cancer. PLoS One 10: e0122495. [Crossref]
72. Anayannis NVJ,
Schlecht NF, Belbin TJ (2015) Epigenetic Mechanisms of Human Papillomavirus-Associated
Head and Neck Cancer. Arch Pathol Lab Med 139: 1373-1378. [Crossref]
73. Wentzensen N,
Vinokurova S, von Knebel Doeberitz M (2004) Systematic review of genomic
integration sites of human papillomavirus genomes in epithelial dysplasia and
invasive cancer of the female lower genital tract. Cancer Res 64:
3878-3884. [Crossref]
74. Akagi K, Li J,
Broutian TR, Padilla-Nash H, Xiao W et al. (2014) Genome-wide analysis of HPV
integration in human cancers reveals recurrent, focal genomic instability. Genome
Res 24: 185-199. [Crossref]
75. Smith CL, Bolton A,
Nguyen G (2010) Genomic and epigenomic instability, fragile sites,
schizophrenia and autism. Curr Genomics 11: 447-469. [Crossref]
76. Cooper DN, Mort M,
Stenson PD, Ball EV, Chuzhanova NA (2010) Methylation- mediated deamination of
5-methylcytosine appears to give rise to mutations causing human inherited
disease in CpNpG trinucleotides, as well as in CpG dinucleotides. Hum
Genomics 4: 406-410. [Crossref]
77. Miranda PM, Silva NNT, Pitol BCV, Silva IDCG, Lima-Filho JL et al. (2013) Persistence
or clearance of human papillomavirus infections in women in Ouro Preto, Brazil.
Biomed Res Int 2013: 578276. [Crossref]
78. Newschaffer CJ, Croen LA, Fallin MD, Hertz-Picciotto I, Nguyen DV et al. (2012) Infant
siblings and the investigation of autism risk factors. J Neurodev Disord
4: 7. [Crossref]
79. Zerbo O, Yoshida C,
Gunderson EP, Dorward K, Croen LA (2015) Interpregnancy Interval and Risk of
Autism Spectrum Disorders. Pediatrics 136: 651-657. [Crossref]
80. Maglennon GA,
McIntosh PB, Doorbar J (2014) Immunosuppression facilitates the reactivation of
latent papillomavirus infections. J Virol 88: 710-716. [Crossref]
81. Shim J, Pérez A,
Symanski E, Nyitray AG (2016) Association Between Serum 25-Hydroxyvitamin D
Level and Human Papillomavirus Cervicovaginal Infection in Women in the United
States. J Infect Dis 213: 1886-1892. [Crossref]
82. Lee BK, Eyles DW,
Magnusson C, Newschaffer CJ, McGrath JJ et al. (2021) Developmental vitamin D
and autism spectrum disorders: findings from the Stockholm Youth Cohort. Mol
Psychiatry 26: 1578-1588. [Crossref]
83. Scheller NM,
Pasternark B, Molgaard-Nielsen D, Svanström H, Hviid A (2017) Quadrivalent HPV
Vaccination and the Risk of Adverse Pregnancy Outcomes. N Engl J Med
376: 1223-1233. [Crossref]
84. Holick MF (2015) Vitamin D and brain health: the need for vitamin D supplementation and sensible sun exposure. J Intern Med 277: 90-93. [Crossref]
85. Heerema-McKenney A (2018) Defense and infection of the human placenta. APMIS 126: 570-588. [Crossref]