Extracellular Matrix Expression in Human Induced Pluripotent Stem Cell-Derived Optic Vesicles
A B S T R A C T
Background: Human tissue/organ development is a complex, highly orchestrated process, regulated in part by the surrounding extracellular matrix (ECM). Every complex tissue, including the retina, has a unique ECM configuration that plays a critical role in cellular differentiation, adhesion, migration, and maturation.
Aim: To characterize ECM expression of human induced pluripotent stem cell-derived optic vesicles (iPSC-OVs).
Methods: A 3- dimensional (3D) in vitro suspension culture system was used to direct differentiation of human induced pluripotent stem cells (iPSCs) into optic vesicles (OVs). Stepwise differentiation of iPSCs into retinal progenitor cells was confirmed by sequential expression of OTX2, SOX1, SIX6, LHX2, PAX6, and CHX10. Expression of ECM genes in iPSC-derived OVs was analyzed by RT2 ProfilerTM PCR Array, whereas immunofluorescence staining was performed to detect ECM proteins in the OVs.
Results: A number of cell adhesion molecules (CAMs) previously reported to be abundantly expressed in iPSCs such as E-cadherin, Intercellular adhesion molecule-1 (ICAM1), Integrin-α L, Integrin-α M, Integrin-α 6 were downregulated while neural and retina specific CAMs including neural cell adhesion molecule 1 (NCAM1), neural plakophilin-related armadillo repeat protein (NPRAP), Integrin-α 1 and Integrin-α 4 were upregulated. Several glycoproteins that have been reported to play key roles during retinogenesis, namely CD44, Tenascin C, Tenascin R, Neurocan, Neuroglycan C, Delta 2 Catenin, Vitronectin, and Reelin were also present.
Conclusion: We have identified an array of ECM proteins that were expressed during retinogenesis. Further characterization of these proteins will lead to a better understanding of retinal development.
Keywords
extracellular matrix, induced pluripotent stem cells, optic vesicles, retina, embryoid body, retinogenesis
Introduction
Induced pluripotent stem cells (iPSCs) are derived from adult somatic cells that have been reprogrammed to become pluripotent. iPSCs share similar properties as embryonic stem cells (ESCs), including the plasticity to differentiate into any somatic cell [1, 2]. Several methods have been used to differentiate pluripotent stem cells into various cell types. One of the most common methods generates uniform and size-controlled 3-dimensional (3D) stem cell aggregates called embryoid bodies (EBs) that are further differentiated into organoids of interest [3, 4]. Studies have shown that 3D EBs allow directed iPSC differentiation into desired cell types in a more efficient and controlled manner by recapitulating the key events that occur during early embryogenesis [5, 6]. EBs help to create a microenvironment that is inducive to cell-cell interactions and endogenous autocrine/paracrine signaling essential for self-renewal and differentiation.
Embryogenesis is a highly orchestrated process consisted of stepwise differentiation of pluripotent stem cells. During this process, embryos also start to secrete laminin and other extracellular matrix (ECM) molecules as early as the two-cell stage by a process called exocytosis [7]. As these embryonic cells further divide, each population of cells differentiates into tissue specific somatic cells. An EB-based differentiation approach has often been applied to induce in vitro retinogenesis of iPSCs, in which the sequential development of neuroectoderm specification, along with eye field and optic vesicle (OV) formation have been observed [8, 9]. ECM is a complex, dynamic network of proteins and carbohydrate molecules within the interstitial space and basement membrane of tissue [10, 11]. It is composed of different macromolecules including glycosylated proteins (proteoglycans), integrins, fibrous proteins and polysaccharides [10, 12, 13]. These molecules constitute the cellular microenvironment that provides structural support and exert influences on cell behaviors in various ways [14-17]. By binding to transmembrane proteins, ECM can regulate the underlying intracellular signaling pathways [18]. ECM can also regulate cell migration and differentiation by virtue of its stiffness and has been shown to influence tissue morphogenesis [19, 20]. In addition, ECM can sequester growth factors thereby influencing cell growth by regulating growth factor bioavailability.
ECM also plays a crucial role in degeneration/regeneration of tissue including retina in various diseases and injuries [21]. Several ECM molecules are upregulated in response to retinal damage. For example, Neuroglycan-C, a brain-specific chondroitin sulfate proteoglycan, has been shown to inhibit optic nerve regeneration after nerve crush injury in rats [22]. Other glycoproteins like Neurocan and CD44 are expressed at the outer interface of abutted retinas and inhibit neurite outgrowth [23]. Tenascin-C is also abundantly expressed after nerve injury which is important for axonal regeneration [24]. ECM remodeling also occurs in retinal diseases like glaucoma and diabetic retinopathy [25, 26].
In this study, we used a 3D in vitro suspension culture system to direct differentiation of human iPSCs into OVs and examined their ECM composition during early events of retinogenesis. Based on the results described here, we report that iPSCs undergoing retinal differentiation in 3D produce retina-specific ECMs. Further characterization of selected iPSC-derived ECM proteins can lead to a better understanding of retinal development and shed light on the potential application of stem cell-derived retinal organoids as a source of autologous ECM materials for retinal repair and regeneration.
Materials and Methods
I Culture and Maintenance of iPSCs
Human iPSC line IMR90-1 was obtained from WiCell (Madison, WI). iPSCs were seeded on Matrigel-coated (Corning, NY) 6 well-plates and maintained in mTeSR1 medium (Stem Cell Technologies, Vancouver, BC, Canada) at 37°C with 5% CO2. Differentiated colonies were identified and removed via visual inspection using a dissecting microscope. Cells were manually passaged using Stempro EZPassage (ThermoFisher Scientific, Waltham, MA) when they reached 80% confluency, where approximately 40 to 50 colonies were seeded onto each well of the Matrigel-coated plates. iPSCs were tested for pluripotency via immunostaining for various pluripotency markers as previously described [27].
II 3D In Vitro Retinal Differentiation
A retinal differentiation protocol was adapted from previous studies with modifications [9, 28]. Briefly, iPSCs were treated with 10 µM Rock inhibitor, Y-27632 (Stem Cell Technologies, Vancouver, Canada) for two hours, rinsed with Phosphate Buffered Saline (PBS), and dissociated into single cells using 0.1% Trypsin for 10 minutes. The cells were washed with PBS and resuspended at a density of 90,000 cells/mL in retinal differentiation medium containing G-MEM (ThermoFisher Scientific, Waltham, MA) with 20% KnockOut Serum Replacement (KSR) (ThermoFisher Scientific, Waltham, MA), 0.2 mM nonessential amino acids, 1 mM pyruvate, 0.1 mM 2-mercaptoethanol, 10 µM Y-27632, 3 µM IWR-1-endo (Wnt inhibitor, Calbiochem, Billerica, MA) and antibiotic/antimycotic (ThermoFisher Scientific). These cells were then assembled into uniform and size-controlled EBs of 9000 cells by aliquoting 100 µL of the cell suspension into each well of the Sumilon PrimeSurface 96-well V-bottomed plates (Sumitomo Bakelite, Novi, MI). After 24 hours, 1 µL of Matrigel at 1% v/v was added to each well. Half the medium in each well was changed to fresh medium without Y-27632 on Day 6. Cells were further cultured until Day 12.
On Day 12, the EBs were collected, washed twice and resuspended in medium containing G-MEM with 20% KSR, 10% Fetal bovine serum (FBS), 1% Matrigel, 0.2 mM nonessential amino acids, 1 mM pyruvate, 0.1 mM 2-mercaptoethanol, and 1X antibiotic/antimycotic cocktail and transferred to 6-well ultra-low attachment plate (Corning, Corning, NY) at a density of 30-40 EBs/well and cultured in suspension. Three µM CHIR 99021, (Wnt agonist, R&D Systems, Minneapolis, MN) and 100 nM smoothened agonist (SAG, R&D Systems, Minneapolis, MN) were added to the medium on Day 15. The medium was changed to neuroretina medium containing DMEM/F12-Glutamax, N2 supplement (ThermoFisher Scientific, Waltham, MA), and 1X antibiotic/antimycotic cocktail on Day 18. Cell differentiation was continued until Day 24.
III Immunofluorescence Staining of EBs During Stepwise Retinal Differentiation
EBs were harvested on Day 8, 12, and 24 during stepwise differentiation and fixed in 4% paraformaldehyde (PFA). After overnight fixation, the EBs were immersed in 15% and 30% sucrose (in PBS) for 2 hours each and embedded in optimal cutting temperature (OCT) compound. Embedded molds were frozen gradually and then sliced into 20 µm sections.
Cryosections were permeabilized with 0.1% Triton X-100 and incubated with antibodies against OTX2, SOX1, SIX6, LHX2, PAX6, or CHX10 to detect markers of retinal development (Table 1). Fluorophore-conjugated secondary antibodies (Molecular Probes, Eugene, OR) were used for immunofluorescence detection. Cell nuclei were counterstained with 4, 6-diamidino-2-phenylindole (DAPI). Fluorescent images were acquired using a Leica TCS SP5 II confocal microscope (Leica Microsystems, Inc. Buffalo Grove, IL) and processed using the Image J software.
Table 1: Primary antibodies used for immunofluorescence
assay.
|
Antibody |
Type |
Source |
Dilution |
|
CD44 |
Mouse monoclonal |
Novus |
1:100 |
|
CHX10 |
Rabbit polyclonal |
Novus |
1:100 |
|
LHX2 |
Goat polyclonal |
Santa Cruz |
1:25 |
|
Neurocan |
Mouse monoclonal |
ABCAM |
1:50 |
|
Neurogycan C |
Rabbit polyclonal |
ABCAM |
1:300 |
|
Pax6 |
Mouse monoclonal |
Developmental hybridoma bank |
1:50 |
|
Sox1 |
Goat polyclonal |
R&D Systems |
1:40 |
|
Six6 |
Rabbit polyclonal |
ABCAM |
1:100 |
|
Tenascin C |
Rabbit polyclonal |
ABCAM |
1:150 |
|
Tenascin R |
Rabbit polyclonal |
Novus |
1:50 |
IV RT2 ProfilerTM PCR Array Assay
Total RNA was extracted from iPSCs (day 0) and iPSCs-derived OVs on day 24 using the RNeasy mini kit (Qiagen, Valencia, CA). RNA was purified with RNase-free DNase (Qiagen). The purified RNA was reverse transcribed to cDNA using the RT First strand kit (Qiagen). cDNA was mixed with the RT SYBR Green ROX qPCR mastermix and aliquoted into the Human Extracellular Matrix and Cell Adhesion Molecules RT2 ProfilerTM PCR Array (Qiagen). Applied Biosystem 7300 RT PCR system (Applied Biosystems, Foster City, CA) was used for thermal cycling: segment 1 – 1 cycle: 95°C for 10 minutes, segment 2 – 40 cycles: 95°C for 15 seconds followed by 60°C for 1 minute. Data were analyzed using the RT2 ProfilerTM PCR Array Data Analysis Webportal (Link) according to the manufacturer’s instructions. Fold changes in gene expression were calculated using the ∆∆Ct method. Expression of house-keeping genes was used to normalize the data and calculate ∆∆Ct values.
V Immunofluorescence Staining of ECM
EBs on Day 24 post-differentiation were fixed and processed for immunofluorescence staining following the general protocol as described above. Antibodies against Neuroglycan C, Neurocan, Tenascin C, CD44, Tenascin R, Reelin, NCAM, Delta 2 Catenin, and Vitronectin were used to detect expression of ECM proteins. Fluorophore-conjugated secondary antibodies were used for immunofluorescence detection. DAPI was used to counterstain cell nuclei. Images were acquired using a confocal microscope as previously described. All images were processed using the Image J software.
VI Statistical Analysis
Each experiment was performed in triplicates. Data were analyzed by Student’s t test and presented as the mean ± standard deviation (SD). A value of p < 0.05 was considered statistically significant.
Results
I In Vitro 3D Differentiation of OVs
We applied a well-established retinal differentiation protocol to direct differentiation of iPSCs into OVs with some modifications [9, 28]. IMR90-1 cells were dissociated into single cells and aggregated to form uniform and size-controlled EBs of 9000 cells/EB (Figure 1A). We observed that IMR90-1 cells underwent stepwise differentiation into retinal progenitors. They expressed neuroectodermal marker OTX2 and SOX1 by Day 8 (Figure 1B) and acquired eye field specification markers, SIX6 and LHX2 by Day 12 (Figure 2). These EBs further developed into OVs as marked by co-expression of neuroretina markers PAX6 and CHX10 (Figure 3). Here we have shown that 3D in vitro differentiation of iPSCs into OVs recapitulates in vivo retinogenesis via sequential expression of the neuroectoderm, eye field and neuroretina markers.
II Expression Profile of ECM Genes in iPSC-Derived OVs
Figures 3 showed that OVs derived from iPSCs in 3D culture expressed markers of the neuroretina. These results suggest that iPSC-derived OVs can be used as a model to study the expression of retina-specific ECM. To characterize the ECM, gene expression profiles of iPSCs and iPSC-derived OVs were analyzed with a commercially available RT2 ProfilerTM PCR Array. iPSCs-derived OVs showed significantly downregulated expression of CAMs including E-cadherin, Intercellular adhesion molecule-1 (ICAM1), Integrin-α L, Integrin-α M, and Integrin-α 6 (Figure 4) and upregulated expression of Neural cell adhesion molecule 1 (NCAM1), Neural plakophilin related armadillo protein (NPRAP), Integrin-α 1 and Integrin-α 4 (Figure 4). Furthermore, ECM expression of glycoproteins Tenascin C, CD44, Vitronectin and Fibronectin I was significantly upregulated in iPSCs-derived OVs (Figure 5).
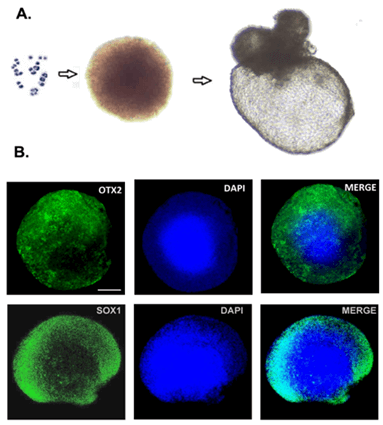
Figure 1: Stepwise differentiation of iPSCs towards a retinal lineage. A)9000 iPSCs were aggregated to form uniform-sized EBs and differentiated towards a retinal lineage. B) Differentiating EBs expressed neuronal marker OTX2 and SOX1 by Day 8 (scale bar=200µm).
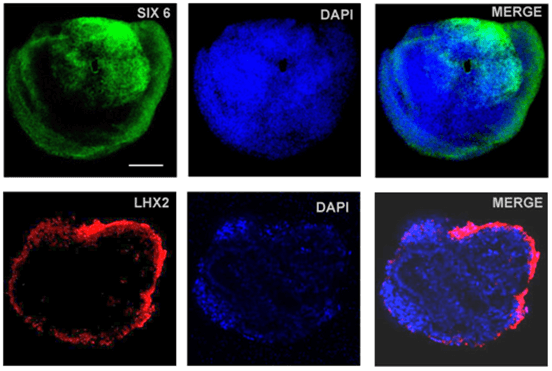
Figure 2: Differentiating EBs expressed eye field specification markers SIX6 and LHX2 by Day 12 (scale bar= 200µm).
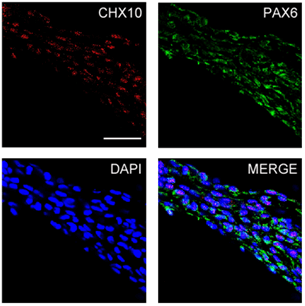
Figure 3: Optic vesicle formation from iPSCs. Developing EBs from iPSCs co-expressed neuronal markers PAX6 and retina progenitor cell marker CHX10 (scale bar=25µm).
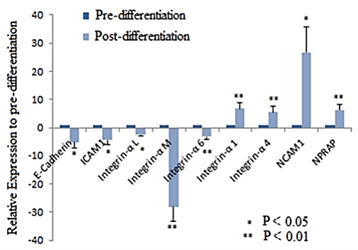
Figure 4: Expression of CAMs and glycoproteins in iPSC-derived OVs on Day 24 of differentiation. iPSC-specific CAMs including E-cadherin, ICAM1, Integrin-α L, Integrin-α M, Integrin-α 6 were downregulated while neural and retina specific CAMs NCAM1, NPRAP, Integrin-α 1 and Integrin-α 4 were upregulated.
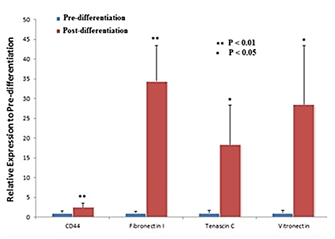
Figure 5: Expression of glycoproteins in iPSC-derived OVs on Day 24 of differentiation. Genes for ECM proteins important for retinogenesis namely Tenascin-C, CD44, Fibronectin I and Vitronectin were upregulated.
III Expression of ECM Proteins in iPSC-Derived OVs
The data from the RT2 ProfilerTM PCR array showed that iPSCs-derived OVs expressed several ECM genes associated with cells of the retinal lineage. Immunofluorescence analysis was performed to further verify the expression and localization of ECM proteins. CD44, a glycoprotein expressed in retinal Müller cells was significantly higher in iPSC-derived OVs as detected by RT-PCR microarray (Figure 4) and via immunofluorescence staining (Figure 7) [29]. Tenascin R, a known modulator of retinal axon growth, was also detected in iPSC-derived OVs (Figure 8) [30]. Additional neural and retinal ECM proteins were found to be expressed in the iPSC-derived OVs, include Reelin, Vitronectin, Delta 2 Catenin, Neuroglycan C, Neurocan, and Tenascin C (Figures 6-8) [31-33].
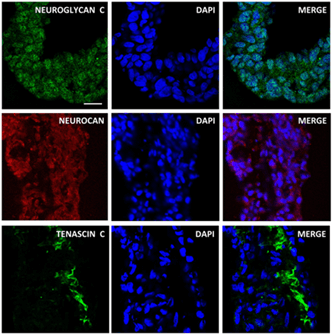
Figure 6: Protein expression of Neuroglycan C, Neurocan, and Tenascin C in iPSC-derived OVs. Neuroglycan C, Neurocan, and Tenascin C were abundantly expressed in the developing iPSC-derived OVs (scale bar=25µm). DAPI (middle column), merge view (right column). These ECM proteins are known to be expressed in the developing eye in vivo.
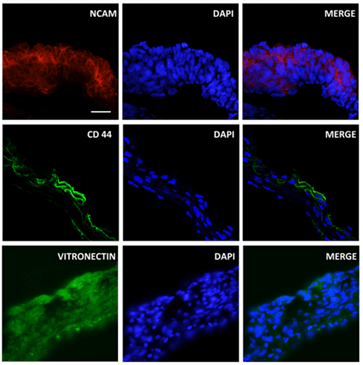
Figure 7: Expression of NCAM, CD44 and Vitronectin in iPSCs-derived OVs. Immunofluorescence analysis showed that OVs expressed NCAM, CD 44 and Vitronectin in the developing OVs (left column). Nuclear staining/DAPI (middle column), merge view (right column), scale bar=50µm.
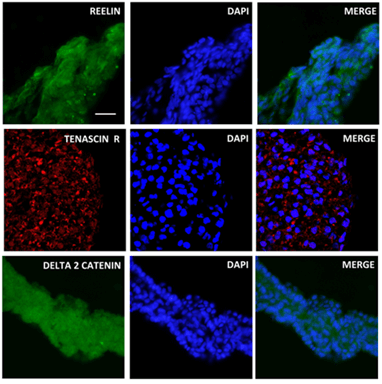
Figure 8: Expression of Reelin, Tenascin R and Delta 2 Catenin in iPSC-derived OVs. Retina abundant proteins like Reelin, Tenascin-R and Delta 2 Catenin were expressed in developing iPSC-derived OVs during early stage retinogenesis. Nuclear staining/DAPI (middle column), merge view (right column), scale bar=50µm.
Discussion
Intracellular and extracellular changes occur during tissue/organ development and provide necessary cues for differentiation to ensure cells can acquire a tissue-specific phenotype [17, 19, 27, 34]. Engineering iPSCs into 3D EBs provides a unique environment that mimics the environment of early embryogenesis [29, 30]. Pluripotent stem cells have shown stepwise acquisition of developmental phenotypes during differentiation towards a defined lineage [3]. During human retinogenesis, the appearance of eye field cells (SIX6+ and LHX2+) in the diencephalon of the anterior neuroectoderm (with SOX1+ and OTX2+ cells) marks the initiation of the neuroretina development (with CHX10+ and PAX6+ retinal progenitor cells). The formation of OVs followed by subsequent invagination of the vesicles results in morphogenesis of the optic cup [8, 31, 32]. Similar sequences of retinal morphogenesis were reported in 3D differentiation of human ESCs and iPSCs in vitro [9, 28]. Even though cellular and phenotypical changes that occur during specification towards a retinal lineage have been well-documented in literature, little is known about the dynamics of ECM formation during retinal development.
In this study, we have demonstrated that iPSC-derived OVs expressed several CAMs/ECMs that have previously been shown to play key roles in retinogenesis in vivo, specifically NCAM1, NPRAP, Tenascin C, CD44, Fibronectin I, and Vitronectin. The iPSCs-derived OVs also expressed Tenascin R, Neurocan, Neuroglycan C, Reelin, Laminin S, and Delta 2 Catenin. Neuroglycan C is a neural tissue specific transmembrane chondroitin sulfate proteoglycan that regulates the formation of the retinal neural network [35, 36]. Another glycoprotein, Tenascin C works in conjunction with Tenascin R to regulate axonal growth during retinogenesis [24, 37]. Furthermore, Tenascin C has also been shown to interact with Neurocan, another glycoprotein that is expressed in neuronal tissue [38, 39]. CD44, another cell surface glycoprotein that was upregulated in this study, is known to be expressed by Müller cells in the retina in vivo [40, 41]. Finally, Fibronectin, and Vitronectin are integral components of the human retina which were also found to be significantly overexpressed in OVs derived from iPSCs [38-41].
The application of biological ECM scaffolds for tissue repair and regeneration is a rapidly advancing interdisciplinary field [33, 42]. A recent study showed that decellularized bovine retinal ECM can support attachment and growth of human retinal progenitor cells [43]. ECM scaffolds derived from CNS tissue have also been reported to support migration and proliferation of neural stem cells [44, 45]. Furthermore, another study demonstrated that ECM scaffolds derived from skin promoted cell engraftment, proliferation and wound healing without scar formation [46, 47]. Many groups have confirmed that tissue-specific ECM promotes maturation and integration when seeded with corresponding cell types [7, 20, 48-54]. These studies indicate that native ECM is a good candidate as a biomaterial for tissue engineering applications.
ECM can be acquired from different sources including human (allogeneic) or animal (xenogeneic) donor tissues [48]. However, it is important to note that animal-derived ECM products pose risks of immunoreactivity and zoonotic disease transmission, whereas human donor tissues are often in short supply [50, 55-60]. Since patient-derived materials are generally preferred for tissue replacement therapy, generating ECM from iPSC-derived organoids could provide autologous materials ideal for tissue repair and regeneration. Using stem cells as a potential source for tissue-specific ECM is a novel approach. There are a few reports on stem cell-derived ECM applications in the literature, including a recent study that described the ability of ECM derived from undifferentiated EBs to support ESC proliferation and differentiation [7]. Several studies have shown that while undergoing chondrogenic differentiation, MSC-derived ECM can induce chondrogenic differentiation of seeded stem cells without exogenous growth factors [61, 62]. These studies have demonstrated the feasibility of obtaining chondrogenic ECM for cartilage regeneration from MSC aggregates differentiating towards a chondrogenic phenotype.
In summary, we have successfully used human iPSCs as a tool to recapitulate early stages of retinal development in this study. We have shown that developing OVs expressed several ECM proteins that are known to play critical roles during retinogenesis in vitro. We have also demonstrated that iPSCs in 3D suspension culture can generate native retina-specific ECM. Future work will include examining the potential of iPSC-derived OVs as an autologous source of ECM materials that can support axonal survival, growth, and guidance for retinal regeneration. Further studies will also be needed to validate the regenerative potency of autologous retina-specific ECM in retinal disease/injury.
Acknowledgement
This work was supported by the U.S. Army Clinical Rehabilitative Medicine Research Program (CRMRP), Military Operational Medicine Research Program (MOMRP) and Oak Ridge Institute for Science and Education (ORISE).
Disclaimers
The opinions or assertions contained herein are the private views of the authors and are not to be construed as official or as reflecting the views of the Department of the Army or the Department of Defense.
Author Contributions
Kaini RR performed data collection, analysis and wrote the manuscript. Rettinger CL performed data analysis and wrote the manuscript. Wang HC and Kaini RR designed the experiments. Wang HC obtained funding, supervised the study, and revised the manuscript. All authors approved the manuscript.
Article Info
Article Type
Research ArticlePublication history
Received: Mon 16, Aug 2021Accepted: Thu 02, Sep 2021
Published: Tue 21, Sep 2021
Copyright
© 2023 Heuy-Ching Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.RGM.2021.02.01
Figures & Tables
Table 1: Primary antibodies used for immunofluorescence
assay.
|
Antibody |
Type |
Source |
Dilution |
|
CD44 |
Mouse monoclonal |
Novus |
1:100 |
|
CHX10 |
Rabbit polyclonal |
Novus |
1:100 |
|
LHX2 |
Goat polyclonal |
Santa Cruz |
1:25 |
|
Neurocan |
Mouse monoclonal |
ABCAM |
1:50 |
|
Neurogycan C |
Rabbit polyclonal |
ABCAM |
1:300 |
|
Pax6 |
Mouse monoclonal |
Developmental hybridoma bank |
1:50 |
|
Sox1 |
Goat polyclonal |
R&D Systems |
1:40 |
|
Six6 |
Rabbit polyclonal |
ABCAM |
1:100 |
|
Tenascin C |
Rabbit polyclonal |
ABCAM |
1:150 |
|
Tenascin R |
Rabbit polyclonal |
Novus |
1:50 |
References
1.
Robinton DA, Daley GQ (2012)
The promise of induced pluripotent stem cells in research and therapy. Nature
481: 295-305. [Crossref]
2.
Yu J, Vodyanik MA, Smuga
Otto K, Antosiewicz Bourget J, Frane JL et al. (2007) Induced pluripotent stem
cell lines derived from human somatic cells. Science 318: 1917-1920. [Crossref]
3.
Ader M, Tanaka EM (2014) Modeling human development in 3D culture.
Curr Opin Cell Biol 31: 23-28. [Crossref]
4.
Clevers H (2016) Modeling
Development and Disease with Organoids. Cell 165: 1586-1597. [Crossref]
5.
Bratt Leal AM, Carpenedo RL,
McDevitt TC (2009) Engineering the
embryoid body microenvironment to direct embryonic stem cell differentiation.
Biotechnol Prog 25: 43-51. [Crossref]
6.
Yodmuang S, Marolt D, Marcos
Campos I, Gadjanski I, Vunjak Novakovic G et al. (2015) Synergistic effects of
hypoxia and morphogenetic factors on early chondrogenic commitment of human
embryonic stem cells in embryoid body culture. Stem Cell Rev Rep 11:
228-241. [Crossref]
7.
Goh SK, Olsen, Banerjee I
(2013) Extracellular matrix aggregates from differentiating embryoid bodies as
a scaffold to support ESC proliferation and differentiation. PLoS One 8:
e61856. [Crossref]
8.
Meyer JS, Howden SE, Wallace
KA, Verhoeven AD, Wright LS et al. (2011) Optic vesicle‐like structures derived from human pluripotent
stem cells facilitate a customized approach to retinal disease treatment. Stem
Cells 29: 1206-1218. [Crossref]
9.
Nakano T, Ando S, Takata N,
Kawada M, Muguruma K et al. (2012) Self-formation of optic cups and storable
stratified neural retina from human ESCs. Cell Stem Cell 10: 771-785. [Crossref]
10.
Frantz C, Stewart KM, Weaver
VM (2010) The extracellular matrix at a glance. J Cell Sci 123:
4195-4200. [Crossref]
11.
Tanzer ML (1985)
Extracellular matrix: extracellular matrix biochemty. Science 227:
289-290. [Crossref]
12.
Halper J, Kjaer M (2014)
Basic components of connective tissues and extracellular matrix: elastin,
fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and
thrombospondins. Adv Exp Med Biol 802: 31-47. [Crossref]
13.
Kular JK, Basu S, Sharma RI
(2014) The extracellular matrix: Structure, composition, age-related
differences, tools for analysis and applications for tissue engineering. J
Tissue Eng 5: 2041731414557112. [Crossref]
14.
Boretti MI, Gooch KJ (2008)
Effect of extracellular matrix and 3D morphogenesis on islet hormone gene
expression by Ngn3-infected mouse pancreatic ductal epithelial cells. Tissue
Eng Part A 14: 1927-1937. [Crossref]
15.
Faulk DM, Johnson SA, Zhang
L, Badylak SF (2014) Role of the
extracellular matrix in whole organ engineering. J Cell Physiol
229: 984-989. [Crossref]
16.
Streuli C (1999)
Extracellular matrix remodelling and cellular differentiation. Curr Opin
Cell Biol 11: 634-640. [Crossref]
17.
Watt FM, Huck WT (2013) Role of the extracellular matrix in regulating
stem cell fate. Nat Rev Mol Cell Biol 14: 467-473. [Crossref]
18.
Vecino E, Heller JP, Veiga
Crespo P, Martin KR, Fawcett JW (2015) Influence of extracellular matrix
components on the expression of integrins and regeneration of adult retinal
ganglion cells. PLoS One 10: e0125250. [Crossref]
19.
Bonnans C, Chou J, Werb Z
(2014) Remodelling the extracellular
matrix in development and disease.
Nat Rev Mol Cell Biol 15: 786-801. [Crossref]
20.
Varshney S, Hunter DD,
Brunken WJ (2015) Extracellular Matrix Components Regulate Cellular Polarity
and Tissue Structure in the Developing and Mature Retina. J Ophthalmic Vis
Res 10: 329-339. [Crossref]
21.
Ren T, van der Merwe Y,
Steketee MB (2015) Developing Extracellular Matrix Technology to Treat Retinal
or Optic Nerve Injury(1,2,3). eNeuro 2: ENEURO.0077-15.2015. [Crossref]
22.
Reinhard J, Renner M,
Wiemann S, Shakoor DA, Stute G et al. (2017) Ischemic injury leads to
extracellular matrix alterations in retina and optic nerve. Sci Rep 7:
43470. [Crossref]
23.
Zhang Y, Kardaszewska AK,
van Veen T, Rauch U, Perez MTR et al. (2004) Integration between abutting
retinas: role of glial structures and associated molecules at the interface. Invest
Ophthalmol Vis Sci 45: 4440-4449. [Crossref]
24.
Besser M, Jagatheaswaran M,
Reinhard J, Schaffelke P, Faissner A (2012) Tenascin C regulates proliferation
and differentiation processes during embryonic retinogenesis and modulates the
de-differentiation capacity of Müller glia by influencing growth factor
responsiveness and the extracellular matrix compartment. Dev Biol 369:
163-176. [Crossref]
25.
Roy S, Amin S, Roy S (2016)
Retinal fibrosis in diabetic retinopathy. Exp Eye Res 142: 71-75.
[Crossref]
26.
Schneider M, Fuchshofer R
(2016) The role of astrocytes in optic nerve head fibrosis in glaucoma. Exp
Eye Res 142: 49-55. [Crossref]
27.
Kaini RR, Shen Gunther J,
Cleland JM, Greene WA, Wang HC et al. (2016) Recombinant Xeno-Free Vitronectin
Supports Self-Renewal and Pluripotency in Protein-Induced Pluripotent Stem
Cells. Tissue Eng Part C Methods. [Crossref]
28.
Sasai Y, Eiraku M, Suga H
(2012) In vitro organogenesis in three dimensions: self-organising stem cells. Development
139: 4111-4121. [Crossref]
29.
Sutha K, Schwartz Z, Wang Y,
Hyzy S, Boyan BD et al. (2015) Osteogenic embryoid body-derived material
induces bone formation in vivo. Sci Rep 5: 9960. [Crossref]
30.
Sepulveda DE, Andrews BA,
Asenjo JA, Papoutsakis ET (2008) Comparative transcriptional analysis of
embryoid body versus two-dimensional differentiation of murine embryonic stem
cells. Tissue Eng Part A 14: 1603-1614. [Crossref]
31.
Chow RL, Lang RA (2001)
Early eye development in vertebrates. Annu Rev Cell Dev Biol 17:
255-296. [Crossref]
32.
Eiraku M, Takata N, Ishibashi H, Kawada M, Sakakura
E et al. (2011) Self-organizing
optic-cup morphogenesis in three-dimensional culture. Nature 472: 51-56.
[Crossref]
33.
O'Brien FJ (2011)
Biomaterials & scaffolds for tissue engineering. materialstoday 14:
88-95.
34.
Hausman RE (2007) Ocular
extracellular matrices in development. Prog Retin Eye Res 26: 162-188. [Crossref]
35.
Escher P, Cottet S, Aono S,
Oohira A, Schorderet DF (2008) Differential neuroglycan C expression during
retinal degeneration in Rpe65-/- mice. Mol Vis 14: 2126-2135. [Crossref]
36.
Inatani M, Tanihara H, Oohira A, Otori Y, Nishida A
et al. (2000) Neuroglycan C, a
neural tissue-specific transmembrane chondroitin sulfate proteoglycan, in
retinal neural network formation. Invest Ophthalmol Vis Sci 41:
4338-4346. [Crossref]
37.
Schumacher S, Stube EM
(2003) Regulated binding of the fibrinogen-like domains of tenascin-R and
tenascin-C to the neural EGF family member CALEB. J Neurochem 87:
1213-1223. [Crossref]
38.
Inatani M, Tanihara H (2002) Proteoglycans in
retina. Prog Retin Eye Res 21: 429-447. [Crossref]
39.
Kwan KM (2014) Coming into
focus: the role of extracellular matrix in vertebrate optic cup morphogenesis. Dev
Dyn 243: 1242-1248. [Crossref]
40.
Chaitin MH, Ankrum MT,
Wortham HS (1996) Distribution of CD44 in the retina during development and the
rds degeneration. Brain Res Dev Brain Res 94: 92-98. [Crossref]
41.
Reinhard J, Joachim SC,
Faissner A (2015) Extracellular matrix remodeling during retinal development. Exp
Eye Res 133: 132-140. [Crossref]
42.
Yi S, Ding F, Gong L, Gu X
(2017) Extracellular Matrix Scaffolds for Tissue Engineering and Regenerative
Medicine. Curr Stem Cell Res Ther 12: 233-246. [Crossref]
43.
Kundu J, Michaelson A,
Talbot K, Baranov P, Young MJ et al. (2016) Decellularized retinal matrix:
Natural platforms for human retinal progenitor cell culture. Acta Biomater
31: 61-70. [Crossref]
44.
Crapo PM, Medberry CJ, Reing
JE, Tottey S, van der Merwe Y et al. (2012) Biologic scaffolds composed of
central nervous system extracellular matrix. Biomaterials 33: 3539-3547. [Crossref]
45.
Crapo PM, Tottey S, Slivka
PF, Badylak SF (2014) Effects of biologic scaffolds on human stem cells and
implications for CNS tissue engineering. Tissue Eng Part A 20: 313-323.
[Crossref]
46.
Wainwright DJ (1995) Use of
an acellular allograft dermal matrix (AlloDerm) in the management of
full-thickness burns. Burns 21: 243-248. [Crossref]
47.
Zhong SP, Zhang YZ, Lim CT
(2010) Tissue scaffolds for skin wound healing and dermal reconstruction. Wiley
Interdiscip Rev Nanomed Nanobiotechnol 2: 510-255. [Crossref]
48.
Choi YC, Choi JS, Kim BS,
Kim JD, Yoon HI et al. (2012) Decellularized extracellular matrix derived from
porcine adipose tissue as a xenogeneic biomaterial for tissue engineering. Tissue
Eng Part C Methods 18: 866-876. [Crossref]
49.
Cigognini D, Lomas A, Kumar
P, Satyam A, English A et al. (2013) Engineering in vitro microenvironments for
cell based therapies and drug discovery. Drug Discov Today 18:
1099-1108. [Crossref]
50.
Eitan Y, Sarig U, Dahan N,
Machluf M (2010) Acellular cardiac extracellular matrix as a scaffold for
tissue engineering: in vitro cell support, remodeling, and biocompatibility. Tissue
Eng Part C Methods 16: 671-683. [Crossref]
51.
Lancaster MA, Knoblich JA
(2014) Organogenesis in a dish: modeling development and disease using organoid
technologies. Science 345: 1247125. [Crossref]
52.
Lin YM, Zhang A, Rippon HJ,
Bismarck A, Bishop AE (2010) Tissue engineering of lung: the effect of
extracellular matrix on the differentiation of embryonic stem cells to
pneumocytes. Tissue Eng Part A 16: 1515-1526. [Crossref]
53.
Turner AEB, Flynn LE (2012)
Design and characterization of tissue-specific extracellular matrix-derived
microcarriers. Tissue Eng Part C Methods 18: 186-197. [Crossref]
54.
Xing Q, Yates K, Tahtinen M,
Shearier E, Qian Z et al. (2015) Decellularization of fibroblast cell sheets
for natural extracellular matrix scaffold preparation. Tissue Eng Part C
Methods 21: 77-87. [Crossref]
55.
Denner J, Mueller NJ (2015)
Preventing transfer of infectious agents. Int J Surg 23: 306-311. [Crossref]
56.
Tang C, Jin C, Du X, Yan C,
Min BH et al. (2014) An autologous bone marrow mesenchymal stem cell-derived
extracellular matrix scaffold applied with bone marrow stimulation for
cartilage repair. Tissue Eng Part A 20: 2455-2462. [Crossref]
57.
Turner NJ, Yates AJ Jr,
Weber DJ, Qureshi IR, Stolz DB et al. (2010) Xenogeneic extracellular matrix as
an inductive scaffold for regeneration of a functioning musculotendinous
junction. Tissue Eng Part A 16: 3309-3317. [Crossref]
58.
Unger C, Skottman H, Blomberg P, Dilber MS, Hovatta
O et al. (2008) Good manufacturing
practice and clinical-grade human embryonic stem cell lines. Hum Mol Genet
17: R48-R53. [Crossref]
59.
Fong AH, Romero López M,
Heylman CM, Keating M, Tran D et al. (2016) Three-dimensional adult cardiac
extracellular matrix promotes maturation of human induced pluripotent stem
cell-derived cardiomyocytes. Tissue Eng Part A 22: 1016-1025. [Crossref]
60.
Pati F, Jang J, Ha DH, Kim
SW, Rhie JW et al. (2014) Printing three-dimensional tissue analogues with
decellularized extracellular matrix bioink. Nat Commun 5: 3935. [Crossref]
61. Cai R, Nakamoto T, Kawazoe N, Chen G (2015) Influence of stepwise chondrogenesis-mimicking 3D extracellular matrix on chondrogenic differentiation of mesenchymal stem cells. Biomaterials 52: 199-207. [Crossref]
62. Mehlhorn AT, Niemeyer P, Kaiser S, Finkenzeller G, Stark GB et al. (2006) Differential expression pattern of extracellular matrix molecules during chondrogenesis of mesenchymal stem cells from bone marrow and adipose tissue. Tissue Eng 12: 2853-2862. [Crossref]