Evidence for Edible Chinese Herbal Medicine as an Alternative Approach for the Treatment of Colorectal Cancer
A B S T R A C T
Colorectal cancer (CRC) is the leading cause of cancer lethality worldwide, inflicting a heavy burden on society. Unfortunately, the complex nature of CRC pathogenesis renders current chemotherapies and targeted therapies unsatisfactory. Conventional medicine generally concentrates on the killing of tumor cells by inducing cell death or activation of the immune system, which often leads to adverse effects or relapsed chemoresistance. On the contrary, Chinese medicine theory pays attention to the patients’ whole inner system and helps to shrink the tumor with consideration of overall body condition. Since numerous Chinese herbal medicines (CHM) are used as food, the edible CHMs as diet resources therapy represent a promising alternative for the treatment of CRC. Recent research has made remarkable progress toward the therapeutic effects of edible CHM for CRC. In this review, experimental CRC models, the holistic characteristic of edible CHM, and recent research on the efficacy and mechanisms of edible CHM as anti-CRC agents are discussed, as well as the safety aspect of edible CHM.
Keywords
Chinese herbal medicine, colorectal cancer, functional food, diet therapy
Introduction
Colorectal cancer (CRC) is a carcinoma, generally an adenocarcinoma, in the colon or rectum [1]. It is amongst the most common cancers worldwide, affecting approximately 3.5 million people with 1.4 million incidences and 694,000 mortalities in 2012 [2]. The treatment of CRC relies on several factors including the size, location, and the stage of the tumor, whether or not it is recurrent, and the overall health status of the patient. During recent decades, various novel therapeutic approaches for CRC have been developed, which led to a steady improvement in the results of treatment. In general, the three most common approaches to treatment are surgery, chemotherapy, and radiotherapy.
A majority of CRC can be cured by removal (endoscopic or surgical approach) of the primary tumor if the disease is in early stages (stages 0 to III according to Tumor-node-metastasis (TNM) classifications [3]. However, for patients at an advanced stage of disease (stage IV), apart from the removal of the tumor, both primary and metastasized if applicable, chemotherapy such as 5-fluorouracil (5-FU), leucovorin, and oxaliplatin combining with targeted therapy are the most commonly used. Initially, some patients (40-50%) respond to the chemotherapy, but eventually, drug resistance (intrinsic or acquired) follows, thereby limiting the clinical efficacy.
Under this situation, many researchers have focused their attention on natural products with anti-CRC activities. As a holistic and combinational approach, the application of Chinese herbal medicines (CHM) in the management of CRC has its unique advantages. Over the past few years, the use of CHM has become increasingly popular among cancer patients worldwide. Correspondingly, there are increasing numbers of pre-clinical and clinical studies conducted to provide evidence for the use of CHM as adjuvant CRC treatment or to provide meaningful information for the development of more effective anti-CRC drugs. Interestingly, many CHMs are consumed for both medicinal and food purposes in the form of traditional food dishes and herbal products. The line between the medicinal and food characteristics of these herbal medicines is often blurred due to the dual uses. Moreover, many Chinese traditional dishes and cuisines may contain one or more types of herbal medicines. Additionally, the concept of diet therapy is widely accepted. Therefore, the introduction of herbal food products that contain CHM as diet therapy would be a promising alternative for CRC management. The present review herein aims to summarize the role of edible CHMs as functional food for diet therapy in CRC management with an emphasis on their underlying mechanisms.
Mechanism of CRC Development
Accumulating evidence shows that CRC is a complex and heterogeneous disease. Three known major molecular groups accounting for the development of CRC have been identified. First, the chromosomal instable group which most commonly features an increase of mutations in specific oncogenes and tumor suppressor genes. Second, the microsatellite instable group, which results from genetic hypermutability caused by DNA mismatch repair genes abnormality. The third group is the CpG island methylation phenotype.
Trans¬formation of normal colon epithelial cells into invasive and metastatic CRC cells requires multiple mutations on the genes. The chromosomal instability pathway accounts for around 70% of sporadic CRC and is prototypical for CRC molecular evolution [4]. In the beginning, a pre-malig¬nant precursor lesion (an adenoma) appear after the loss of function mutation of genes in the Wnt pathway APC or CTNNB [5]. Subsequently, mutations of KRAS, TP53, and SMAD4 occur and speed conversion into an invasive and metastatic phenotype (Figure 1) (Cancer Genome Atlas, 2012) [6].
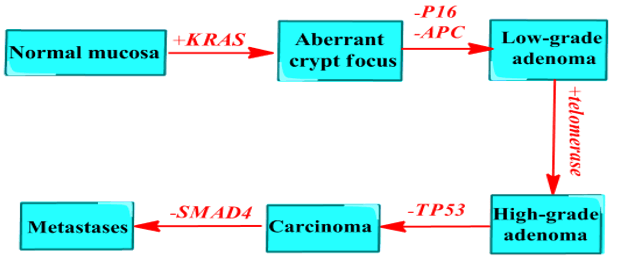
Figure 1: Chromosomal instability pathway in CRC development. The sequence starts from aberrant crypt focus formation often with KRAS activation mutations featuring slight disturbances of crypt structure and in some cases, epithelial dysplasia. Thereafter, adenoma, which is small and generally tubular and has confined cytonuclear and characteristics of dysplasia, follows with frequent inactivation mutations of APC. In the next stage, telomerase is stimulated, and the lesions generally display large, villous architecture and cytonuclear properties of advanced dysplasia. The development of invasive carcinoma is commonly followed by TP53 mutations. Finally, further mutations such as SMAD4 inactivation promote progression towards metastasis.
Edible CHM for CRC Treatment
I Traditional Chinese Medicine Perspective of CRC
Through the perspective of traditional Chinese medicine (TCM) theory, a tumor is not only the morphological changes but also, and more importantly, the functional changes of specific tissues or organs [7]. In TCM, the tumor results from the stagnation of toxin and heat, obstruction of phlegm/dampness, Qi stagnation and blood stasis, and imbalanced yin and yang in viscera and bowels. CRC is caused by the accumulation of toxins. It results from an imbalance of the body with deficient of ‘qi’ and extra toxic fluids and ‘heat’. This situation is further exasperated by a weakened spleen and kidneys, leading to the flow and accumulation of toxins into the intestine. ‘Qi’ deficiency is regarded as the major driving force leading to CRC. TCM discriminate malignant bowel blockage from that of benign. In malignant blockage, there exists both mechanical blockage of the bowel and hindrance of ‘qi’, blood stasis and aggregation of toxins. These issues are thought to have a serious influence on surgical outcomes and needed to be fixed to obtain uncomplicated surgery [8].
In China, CHM has been used for treating cancers for several thousand years. The rationale for using herbal medicine in the treatment of CRC is based upon specific properties of the herbal medicine. Therefore, herbs with properties to accelerate circulation, eradicate blood stasis, clear toxins and heat, energize the spleen and kidneys and most importantly, replenish ‘qi’ are becoming popular as major remedies [8]. The herbal preparation often consists of many ingredients, which, based upon the clinical experiences, can act synergistically to exert its effects in various ways. Firstly, it can alleviate the damage caused by chemo/radiotherapy against the normal cells and tissues in the body. Secondly, it can strengthen the efficacy of chemo/radiotherapy. Thirdly, it can ameliorate inflammatory and infectious complications in normal tissues adjoining the tumor. Fourth, it can boost immunity and body resistance. Fifth, it can make better the patients’ overall condition and quality of life. Last but not least, it can extend the life expectancy of the patients in an advanced stage of cancer [9].
II In Vitro Evidence of Edible CHM for CRC
CHM and their active components have been shown to exhibit cytotoxic activities in various human CRC cell lines with HT-29, LoVo, CaCo-2, SW-480 and -620, HCT-8 and -116, and colo-205 being the most common ones (Table 1).
Among the CHM used for both the medicinal and food purposes, Coptis chinensis (Huanglian), Curcuma longa (Jianghuang), Hedyotis diffusa (Baihuasheshecao), Panax notoginseng (Sanqi), Panax quinquefolius, Salvia miltiorrhiza (Danshen), Scutellaria baicalensis (Huangqin), and Zingiber officinale (Shengjiang) are the most intensely studied (Table 1). The images of these CHMs were shown in (Figure 2), and some of their anti-CRC active compounds are shown in (Figure 3). It has been reported that extracts from C. chinensis could inhibit the proliferation of HCT-116 cells by suppressing cyclin B1 expression and inhibiting CDC2 kinase activity and induce apoptosis in SUN-C4 cells [10, 11].
Berberine, an isoquinoline alkaloid, isolated from C. chinensis has been reported to stimulate apoptosis through inducing ROS stress and activation of JNK/p38 MAPK and FasL in SW-620 cells, and to inhibit invasion and metastasis of SW-620 and LoVo cells via cyclooxygenase-2 (COX-2)/prostaglandin E2 (PGE2) mediated JAK2/STAT3 signaling pathway [11, 12]. Moreover, curcumin, a polyphenol, isolated from C. longa has been reported to induce apoptosis in HT-29, colo-205, and LoVo cells, and to inhibit migration and invasion in colo-205 cells by inhibiting NF-κB/p65 and down-regulating COX-2 [13, 14, 15].
Table 1: In vitro study of edible CHM for CRC treatment.
|
Species/ Formulation |
Extract/ compound |
Cell lines |
Pharmacodynamic indicators |
Refs |
|||
|
C. chinensis |
Methanol extracts |
SNU-C4 |
¯Viabilities; ↑apoptosis populations; ↑Bax; ↑caspase3; ¯Bcl-2; |
(Kim et al. 2004) [56] |
|||
|
|
Water extracts |
HCT-116 |
¯Viabilities; ¯colony formation; ↑G2 phase population; ¯cyclin B1; ¯cdc2 kinase activity; |
(Li et al. 2000) [10] |
|||
|
|
Berberine |
SW-620 |
↓Viabilities;↑ caspase 3 and 8 activation; ↑PARP cleavage; ¯Bid, c-IAP1, Bcl-2, Bcl-XL; ↑p-JNK, p-p38; ↑ROS; ↑p-c-jun, FasL, t-Bid; |
(Hsu, Hsieh, et al. 2007) [20] |
|||
|
C. longa |
Curcuminoids |
5 primary cells |
¯Viabilities; ↑apoptosis population; ¯mitochondrial membrane potential (MMP); |
(Hsu, Weng, et al. 2007) [21] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯COX-2; |
(Du et al. 2006) [57] |
|||
|
|
Curcumin |
Colo-205 |
¯Viabilities; ¯NF-κB p65, COX-2; ↑COX-1; ¯ MMP2/9; ¯invasion ability; |
(Su, Chen, et al. 2006) [58] |
|||
|
|
Curcumin |
Caco-2, HT-29 |
¯EGRF expression; ¯ERK and Elk-1 activity; ¯Egr-1 expression and activity; |
(Chen, Xu, and Johnson 2006) [59] |
|||
|
|
Curcumin |
LoVo |
¯Viabilities; ↑LDH release; ↑apoptosis population; ↑caspase-3 and 9 activation; ¯MMP; ↑cytochrome c release, ↑Bax, p53; ¯Bcl-2, survivin; |
(Guo et al. 2013) [15] |
|||
|
|
Curcumin |
Colo-205 |
¯Viabilities; ↑apoptosis population; ↑ROS, Ca2+; ¯MMP; ↑caspase 3 activity; ↑Bax, cytochrome C, p53, p21; ¯Bcl-2; |
(Su, Lin, et al. 2006) [14] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯COX-2; |
(Goel, Boland, and Chauhan 2001) [60] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯apoptosis population; ¯pAkt, COX-2; ↑p-AMPK; ↑G1 population; |
(Lee et al. 2009) [13] |
|||
|
G. lucidum |
Ganoderic acid Me |
HCT-116 |
¯Viabilities; ↑apoptosis population; ↑p53, Bax; ¯MMP; ↑cytochrome c release; ↑caspase 3 activity; |
(Zhou et al. 2011) [46] |
|||
|
|
Triterpenoids |
SW-480 |
¯Cell number; ¯DNA synthesis; |
(Xie et al. 2006) [30] |
|||
|
H. diffusa |
Ethanol extracts |
HT-29 |
¯Viabilities; ↑apoptosis population; ↑DNA fragmentation; ¯MMP; ↑Bax/Bcl-2; ↑caspase 9 and 3 activation; |
(Lin et al. 2010) [61] |
|||
|
|
Ethanol extracts |
HCT-8 |
¯Viabilities; ¯P-glycoprotein, ABC subfamily G member 2; |
(Li, Wang, et al. 2015) [16] |
|||
|
|
Ethanol extracts |
HT-29 |
¯Viabilities; ↑apoptosis population; ¯p-Stat3; ¯Cyclin D1, CDK4, Bcl-2; ↑p21, Bax; |
(Lin et al. 2015) [17] |
|||
|
|
Ursolic acid |
HT29 |
¯Viabilities; ↑apoptosis population; ¯p-EGFR, p-ERK1/2, p-p38, and p-JNK; ¯Bcl-2, Bcl-xl; ↑caspase 3 and 9 activity; |
(Shan et al. 2009) [62] |
|||
|
|
Ursolic acid |
SW-480, LoVo |
¯Viabilities; ¯clone formation; ¯migration ability; ¯MMP9; CDH1 expression; ¯p-Akt, p-ERK; ¯COX-2, PGE2; ↑NF-κB and p300 translocation; ↑apoptosis population; ↑PARP; ↑caspase 3 and 9 cleavage; ↑cytochrome c release; |
(Wang, Liu, et al. 2013) [36] |
|||
|
P. notoginseng |
Panaxadiol |
HCT-116 |
¯Viabilities; ↑apoptosis population; ↑G1 population; |
(Li et al. 2009) [20] |
|||
|
|
N-butanol fraction- |
HCT-116 |
¯Viabilities; ↑apoptosis population; |
(Wang et al. 2007) [25] |
|||
|
|
N-butanol fraction |
SW-480 |
¯Viabilities; ¯DNA synthesis; ↑apoptosis population; ↑S, G2 phase population; |
(Wang, Xie, et al. 2009) [18] |
|||
|
P. quinquefolius |
Steamed extracts |
HCT-116, SW-480 |
↑Apoptosis population; ¯MMP; ROS; ↑NF-κB activation; |
(Li et al. 2010) [63] |
|||
|
|
Ginseng extracts |
HCT-116 |
¯Viabilities; affect ephrin receptor pathway; |
(Luo et al. 2008) [64] |
|||
|
|
Ginsenoside Rh2 |
HCT-116, SW-480 |
¯Viabilities; ↑apoptosis population; ↑ROS; NF-κB activation; ↑Bax, Bad; ¯Bcl-2, Bcl-XL; ↑cytoplasmic vacuoles; |
(Li et al. 2011) [65] |
|||
|
|
Ginsenoside Rg3 |
SW-480 |
¯Migration ability; ¯NF-κB activity; ¯MMP-9, COX-2; |
(Song et al. 2015) [66] |
|||
|
|
Ginsenoside Rg3 |
HCT-116 |
¯Viabilities; ¯colony formation; ¯β-catenin nuclear translocation; ¯PNCA; |
(He et al. 2011) [22] |
|||
|
S. miltiorrhiza |
Cryptotanshinone |
SW-480, HCT-116, LoVo |
↓Viabilities; ↑apoptosis population; ↓anchorage independent growth; ↑caspase cleavage; ↓Bcl-2; ↓p-Stat3; ↓pEGFR; ↑p21 levels; ↓CyclinD1, survivin; |
(Li, Saud, et al. 2015) [24] |
|||
|
|
Tanshinone IIA |
Colo-205 |
↓Viabilities, ↑apoptosis population; ↑p53, p21; ↑cytochrome c release; ↑Fas; ↑caspase 8 and 3 cleavage; |
(Su et al. 2008) [58] |
|||
|
Z. officinale |
Ethanol extracts |
HCT-116, HT-29 |
↓Cell viability; ↑apoptosis population; ↑G1 phase population; |
(Abdullah et al. 2010) [27] |
|||
|
|
6-gingerol |
LoVo |
¯Viabilities; ¯cyclin A, B1, CDK1; ↑p27, p21; ↑ROS, p-p53; ↑G2 phase population; |
(Lin, Lin, and Tsay 2012) [29] |
|||
|
|
6-Gingerol |
HCT-116, HT-29 |
↓Viabilities; ↑apoptosis population; ↑G1 phase population; ↓cyclin D1; ↑nonsteroidal anti-inflammatory drug-activated gene-1; ↓β-catenin translocation; ↑PKCε, GSK-3β activation; |
(Lee, Cekanova, and Baek 2008) [28] |
|||

Figure 2: Images of the medicinal parts of the Chinese herbal medicine used for both medicinal and food purposes.
Clinically, H. diffusa has been used as a main component in some Chinese medicine formulas for CRC treatment. Ethanol extracts of H. diffusa have been shown to inhibit the proliferation of 5-FU resistant HCT-8 cells via downregulation of the expression of p-glycoprotein and ATP-binding cassette subfamily G member 2, and to inhibit proliferation and induce apoptosis in HT-29 cells through inactivation of IL-6-inducible Stat3 pathway [16, 17]. Furthermore, the n-butanol fraction of P. notoginseng has been shown to inhibit proliferation and induce apoptosis in SW-480 cells, while potentiates the cytotoxic effects of 5-FU against HCT-116 cells [18, 19]. Interestingly, panaxadiol from P. notoginseng potentiated the cytotoxic effects of 5-FU against HCT-116 cells [20]. Moreover, ginsenoside Rg3 from P. quinquefolius or P. ginseng inhibited SW-480 cell migration via inhibition of NF-κB, and exerted antiproliferative effects on HCT-116 cells by down-regulating Wnt/β-catenin signaling [21, 22].
Tanshinone IIA, a diterpene quinone, isolated from S. miltiorrhiza could inhibit the growth and induce apoptosis in colo-205 cells [23]. Cryptotanshinone, another compound from S. miltiorrhiza, could suppress the growth and induce apoptosis in SW-480, HCT-116, and LoVo cells via the inhibition of Stat3 [24]. With regard to S. baicalensis, an aglycone-rich fraction from this herb has been shown to stimulate apoptosis in HCT-116 and HT-29 cells through the mitochondrial pathway [25]. Wogonin, a mono-flavonoid from S. baicalensis, could retard the proliferation in HCT-116 cells by Wnt/catenin signaling pathway and inactivation of CDK8 [26].
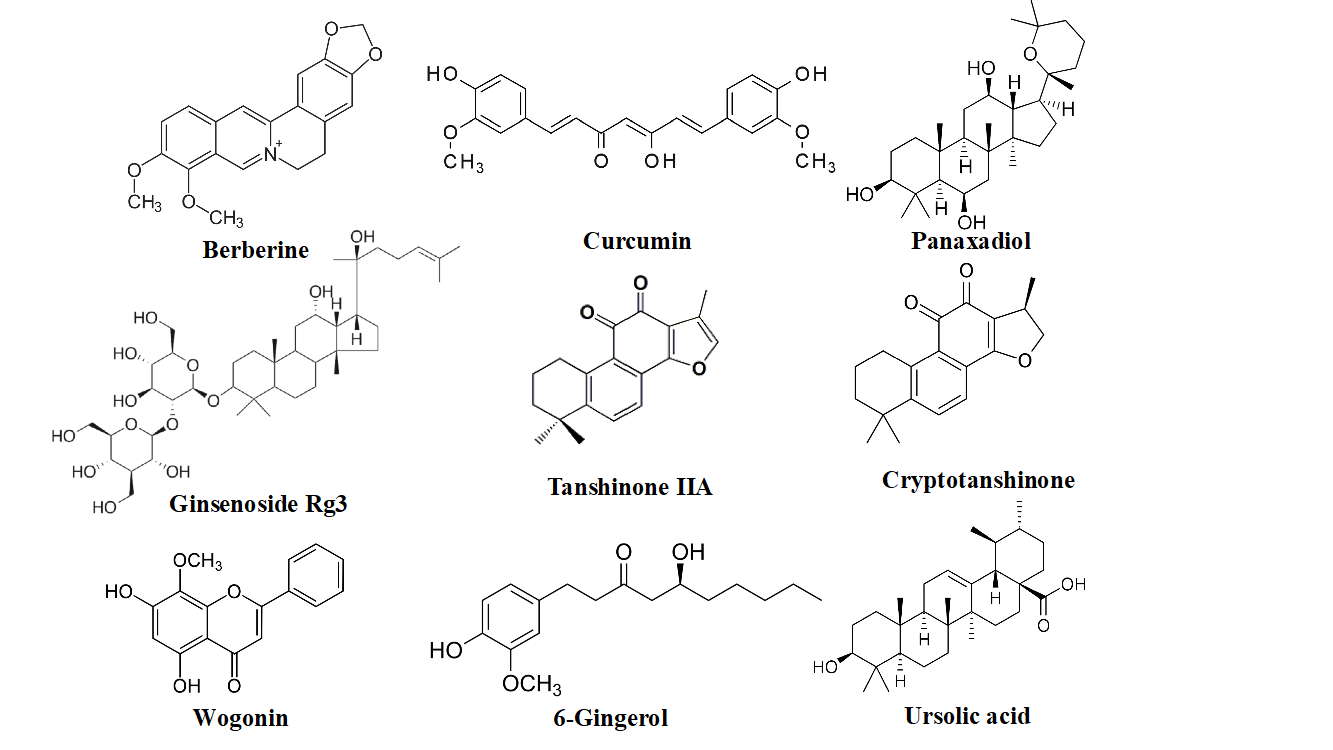
Figure 3: Chemical structures of the representative anti-CRC compounds from edible Chinese herbal medicine.
The other Chinese herb Z. officinale has been used as a TCM for centuries. The ethanol extracts of Z. officinale have been reported to trigger apoptosis and G1-phase arrest in both HCT-116 and HT-29 cells [27]. As a major pharmacologically active component of Z. officinale, 6-gingerol has also been reported to induce apoptosis and G1-phase arrest against HCT-116 as well as HT-29 cells [28]. Interestingly, in LoVo cells, 6-gingerol inhibited the proliferation of the cells via G2-phase arrest [29]. These results indicated that 6-gingerol could inhibit the proliferation of different human CRC cells through different mechanisms.
III In Vivo Evidence of Edible CHM for CRC
Carcinogenesis models are useful to understand the nature of cancer development and explore different approaches to impede the process under in vivo conditions. In this context, 1,2-dimethylhydrazine (DMH)- or azoxymethane (AOM)-initiated and dextran sulfate sodium (DSS)-induced mice/rat experimental colon carcinogenesis models are excellent models of CRC and have been widely used in herbal medicine studies. Moreover, genetic models of intestinal carcinogenesis (APC min) and xenograft models have also been extensively used to determine the chemo preventive and chemotherapeutic effects of herbal medicines or their active compounds (Table 2).
Table 2: In vivo study of Chinese herbal medicine for CRC treatment.
|
Species |
Extract/compound |
In vivo model |
Pharmacodynamic indicators |
Refs |
|
C. chinensis |
Berberine |
SW-620 and LoVo xenograft and lung metastasis |
↓Tumor volume; ↓tumor foci number; |
(Liu et al. 2015) [12] |
|
|
Berberine |
DMH and DSS-induced tumor |
↓Tumor number; ↓tumor incidence; |
(Wu et al. 2012) [67] |
|
|
Berberine |
APC Min/+ mice; |
¯Polyps number and size; ↓cyclin D1, c-Myc; |
(Zhang et al. 2013) [34] |
|
|
Berberine |
AOM and DSS-induced tumor |
↓Tumor number and size; ↓Ki-67, COX-2; ↑AMPK; ↓p-mTOR, p-p65; ↑caspase 3 cleavage; |
(Li, Hua, et al. 2015) [68] |
|
C. longa |
Curcumin |
C-26 xenograft; zebrafish; |
¯Vascular formation; ¯tumor volume; |
(Gou et al. 2011) [69] |
|
G. lucidum |
Triterpenes |
HT-29 xenograft |
↓Tumor volume; ↓Ki-67; ↑Beclin-1, LC-3; |
(Thyagarajan et al. 2010) [70] |
|
H. diffusa |
Ethanol extract |
HT-29 xenograft |
↓Tumor volume; ↓intratumoral microvessel density; ↓SHH, PTCH-1, SMO, Gli-1; ¯VEGF-A, VEGFR2; |
(Lin et al. 2013) [71] |
|
|
Ethanol extract |
HT-29 xenograft |
↓Tumor volume and weight; ↑apoptosis population; ↓PCNA; ↓p-Stat3; ↑Bax, p21; ↓Bcl-2, cyclin D1, CDK4; |
(Cai et al. 2012) [72] |
|
|
Aqueous extract |
HCT-116 xenograft |
↓Tumor volume and weight; ↑p-AMPK ;↓mTORC1; ↑p53 activation; |
(Lu et al. 2016) [38] |
|
P. quinquefolius |
protopanaxadiol |
HCT-116 xenograft |
↓Tumor size; ↓PCNA; |
(Gao et al. 2013) [12] |
|
R. Astragali |
Astragalus saponins |
HT-29 xenograft |
↓Tumor volume; ↓p-Akt, p-mTOR, VEGF, VEGFR1, VEGFR2; ↓COX-2; |
(Law et al. 2012) [73] |
|
|
Astragalus saponins |
HT-29 xenograft |
↓Tumor volume and weight; ↓PCNA; |
(Tin et al. [39] 2007) |
|
|
Astragalus saponins |
HCT-116 xenograft |
↓Tumor volume and weight; ↓PCNA; ↓VEGF; |
(Auyeung, Law, and Ko 2014) [41] |
|
S. miltiorrhiza |
Tanshinone IIA |
Colo-205 xenograft; |
↓Tumor volume; ↓P-glycoprotein, LC-3 II; ↓VEGF, NF-κB p65, and MMP 7; |
(Su 2012) [74] |
|
S. baicalensis |
Baicalein |
AOM and DSS-induced tumor |
↓Tumor number; |
(Kim et al. 2013) [32] |
|
|
Baicalein |
HCT-116 xenograft |
↓Tumor volume; |
(Wang et al. 2015) |
|
|
Baicalin |
HCT-116 xenograft |
↓Tumor volume; ↑survival rate; |
(Yang et al. 2013) [75] |
|
Z. officinale |
6-gingerol |
HCT-116 xenograft |
↓ tumor volume; ↓ leukotriene B4; ↓leukotriene A4 hydrolase; |
(Jeong et al. 2009) [76] |
Using the DMH or AOM and DSS-induced CRC model, berberine from C. chinensis has been shown to inhibit colon tumorigenesis by AMP-activated protein kinase signaling pathways [30, 31]. Moreover, baicalein (derived from S. baicalensis) has also been reported to inhibit CRC progression in DMH/AOM-DSS-induced model [32, 33]. It has also been shown that berberine could attenuate intestinal polyps’ growth in APC Min mice [34].
The human CRC cell xenograft model is probably the most popular for the study of the therapeutic effects of CHM. In particular, baicalin has been shown to inhibit HCT-116 xenograft tumor growth [35]. Additionally, baicalein inhibited the growth of orthotopic xenograft tumors derived from HCT-116 cells deficient in a mismatch repair gene [36]. Furthermore, it is illustrated that H. diffusa ethanol extracts could inhibit HT-29 xenograft tumor growth via the inhibition of Stat3 signaling pathway, while inhibiting HCT-116 xenograft tumor growth via the activation of AMP-activated protein kinase signaling pathway [37, 38]. Moreover, Astragalus saponins have been reported to inhibit the growth and angiogenesis of HT-29 xenograft tumors [39, 40]. Furthermore, in HCT-116 xenograft tumor, Astragalus saponins have also been shown to inhibit tumor growth and proangiogenic factors generation [41].
IV Clinical Evidence of Edible CHM for CRC
It has been reported that compared with conventional therapy alone, CRC patients receiving herbal medicine and vitamins combined with conventional therapy could decrease the risk of death in stage I by 95%, stage II by 64%, stage III by 29%, and stage IV by 75% [42]. Oral administration of berberine (300 mg, thrice per day) for 6 months significantly decreased the polyp size of familial adenomatous polyposis patients, together with the cyclin D1 expression in the polyp [34]. A cohort study showed that the combined treatment of TCM and western medicine had significant clinical value and potential for reducing the relapse or metastasis rate in stage II and III of CRC after a conventional radical operation [43]. Moreover, a preliminary double-blind, randomized clinical trial indicated that the administration of aged garlic extracts for 12 months significantly decreased the size and number of colonic adenomas in CRC patients [44].
Furthermore, PHY906, a four-herb Chinese medicine formula consists mainly of edible CHM described 1800 years ago, enhanced the anti-tumor effects of the chemotherapy in patients with advanced CRC, as shown in a phase I/II clinical trial (Farrell and Kummar, 2003) [9]. Recently, a systematic review suggested that compared with chemotherapy treatment alone, CHM as adjunctive therapy with chemotherapy had a significant efficacy in terms of prolonging survival, enhancing tumor response, improving quality of life, strengthening the immune system, and alleviating acute adverse effects [45].
V Safety Concerns of Edible CHM
It is generally believed that edible plants, such as crops and vegetables are definitely safe. In reality, it is not always as expected. Toxins may inherently be present in edible plants, for instance, potatoes (Solanum tuberosum) contain steroids alkaloids which are toxic and even teratogenic for humans, Lupinus albus contains up to 5% quinolizidine alkaloids which are also toxic [46]. Although bioactive agents or extracts from edible plants are relatively safe, attention should be paid to the safety of some edible plants when taken frequently. In general, the active constituents in herbal supplements should be isolated before consumption. This procedure ensures the separation of the plant’s products with desired therapeutic benefits from those which are not and even toxic. A case in point is Cinnamomum aromaticum, which is widely used to obtain the spice ‘cinnamon’ [47]. Cinnamon possessed many beneficial health effects, including anti-inflammation, anti-microbial, blood glucose control, reducing cardiovascular disease, boosting cognitive function and reducing risk of colonic cancer [48]. Moreover, C. aromaticum also exhibited an anti-diabetic effect [49]. However, C. aromaticum contains large amounts of coumarins that exert strong anticoagulant, carcinogenic and hepato-toxic properties [50]. Consequently, regular consumption of C. aromaticum in large quantities may be harmful [51]. In addition, the lack of strict quality control over the consumption of herbal supplements also draws our attention to the safe use of edible plants. Strict quality control procedures are imposed during the manufacture of conventional drugs to guarantee the consistency of active ingredients in all samples. However, rigorous quality control procedures are not well followed in the preparation of herbal supplements. Moreover, in some extreme occasions, poisonous plants were regarded as genuine herbs. For example, Kelp (seaweed) tablets, a widely available dietary supplement in health food stores rich in vitamins and minerals, contain varied content of iodine and some seaweed dishes may exceed daily tolerable intake of 1100 mg [52].
Another safety concern when consuming edible CHM is herb-drug pharmacokinetic interaction. In most cases, DN patients are simultaneously using conventional medicine and traditional herbal medicine with the latter as an adjuvant. Comparatively, modern drugs have their merits in specifically targeting disease-related molecules through definite pathways; whereas TCM edible plants have the superiority that their multi-components may exert a synergetic effect and benefit the whole internal milieu of patients, raising the possibility that the combinational administration of TCM edible plants and modern drugs may provide better therapeutic effects on diseases, especially chronic and comprehensive diseases like DN. However, after co-administration, edible plants may reduce the permeability of the conventional drug in the intestinal tract and may also affect its metabolism in the liver and cause hypoglycemia. Research by Puranik and colleagues (2011) found that the technology-based supercritical extract of Cassia auriculata caused a significant reduction in absorption of metformin, indicating the need to include pharmacokinetic herb-drug interaction studies to avoid unexpected side effects [53]. Another case is the commonly used herb St. John’s wort. Hyperforin and hypericin, the active component of St. John’s wort, induce CYP3A4 and P-glycoprotein, respectively. Thus, it is likely that drugs that are metabolized by CYP3A4 and P-glycoprotein will interact with St. John’s wort [54, 55]. Therefore, physicians and the patients need to pay crucial attention to potential adverse herb-drug interactions when TCM edible plants are used as an adjuvant in disease management.
Conclusion
The above described in vitro, in vivo, and clinical results strengthen the fact that edible CHM can interfere with numerous molecular pathways related to cancer initiation and progression. These pieces of evidence undoubtedly highlight the application of edible CHM as novel chemo preventive and chemotherapeutic agents for CRC intervention. In future studies of edible CHM, various molecular mechanisms and targets for tumor growth inhibition, apoptosis, anti-angiogenesis, and particularly metastasis still remain to be resolved. Currently, clinical trials with edible CHM conducted for CRC treatment are very limited. Therefore, in order to take full advantage of edible CHM, much more well-designed clinical trials are necessary for further evaluation of the safety and efficacy of edible CHM against CRC.
Acknowledgements
This study was supported by grants from Seed Funding Programme for Basic Research from HKU (Project No. 201611159232); the Innovation and Technology Support Programme (Project code: UIM/321); and the Government of Hong Kong Special Administrative Region. The founders had no role in the design, analysis or writing of this article. All authors contributed to collecting data for the article, discussion of the content, writing and editing of the manuscript before submission. The authors have no conflicts of interest to declare.
Article Info
Article Type
Review ArticlePublication history
Received: Fri 15, May 2020Accepted: Sat 06, Jun 2020
Published: Fri 26, Jun 2020
Copyright
© 2023 Kalin Yan-Bo ZHANG. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.COR.2020.06.08
Figures & Tables
Table 1: In vitro study of edible CHM for CRC treatment.
|
Species/ Formulation |
Extract/ compound |
Cell lines |
Pharmacodynamic indicators |
Refs |
|||
|
C. chinensis |
Methanol extracts |
SNU-C4 |
¯Viabilities; ↑apoptosis populations; ↑Bax; ↑caspase3; ¯Bcl-2; |
(Kim et al. 2004) [56] |
|||
|
|
Water extracts |
HCT-116 |
¯Viabilities; ¯colony formation; ↑G2 phase population; ¯cyclin B1; ¯cdc2 kinase activity; |
(Li et al. 2000) [10] |
|||
|
|
Berberine |
SW-620 |
↓Viabilities;↑ caspase 3 and 8 activation; ↑PARP cleavage; ¯Bid, c-IAP1, Bcl-2, Bcl-XL; ↑p-JNK, p-p38; ↑ROS; ↑p-c-jun, FasL, t-Bid; |
(Hsu, Hsieh, et al. 2007) [20] |
|||
|
C. longa |
Curcuminoids |
5 primary cells |
¯Viabilities; ↑apoptosis population; ¯mitochondrial membrane potential (MMP); |
(Hsu, Weng, et al. 2007) [21] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯COX-2; |
(Du et al. 2006) [57] |
|||
|
|
Curcumin |
Colo-205 |
¯Viabilities; ¯NF-κB p65, COX-2; ↑COX-1; ¯ MMP2/9; ¯invasion ability; |
(Su, Chen, et al. 2006) [58] |
|||
|
|
Curcumin |
Caco-2, HT-29 |
¯EGRF expression; ¯ERK and Elk-1 activity; ¯Egr-1 expression and activity; |
(Chen, Xu, and Johnson 2006) [59] |
|||
|
|
Curcumin |
LoVo |
¯Viabilities; ↑LDH release; ↑apoptosis population; ↑caspase-3 and 9 activation; ¯MMP; ↑cytochrome c release, ↑Bax, p53; ¯Bcl-2, survivin; |
(Guo et al. 2013) [15] |
|||
|
|
Curcumin |
Colo-205 |
¯Viabilities; ↑apoptosis population; ↑ROS, Ca2+; ¯MMP; ↑caspase 3 activity; ↑Bax, cytochrome C, p53, p21; ¯Bcl-2; |
(Su, Lin, et al. 2006) [14] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯COX-2; |
(Goel, Boland, and Chauhan 2001) [60] |
|||
|
|
Curcumin |
HT-29 |
¯Viabilities; ¯apoptosis population; ¯pAkt, COX-2; ↑p-AMPK; ↑G1 population; |
(Lee et al. 2009) [13] |
|||
|
G. lucidum |
Ganoderic acid Me |
HCT-116 |
¯Viabilities; ↑apoptosis population; ↑p53, Bax; ¯MMP; ↑cytochrome c release; ↑caspase 3 activity; |
(Zhou et al. 2011) [46] |
|||
|
|
Triterpenoids |
SW-480 |
¯Cell number; ¯DNA synthesis; |
(Xie et al. 2006) [30] |
|||
|
H. diffusa |
Ethanol extracts |
HT-29 |
¯Viabilities; ↑apoptosis population; ↑DNA fragmentation; ¯MMP; ↑Bax/Bcl-2; ↑caspase 9 and 3 activation; |
(Lin et al. 2010) [61] |
|||
|
|
Ethanol extracts |
HCT-8 |
¯Viabilities; ¯P-glycoprotein, ABC subfamily G member 2; |
(Li, Wang, et al. 2015) [16] |
|||
|
|
Ethanol extracts |
HT-29 |
¯Viabilities; ↑apoptosis population; ¯p-Stat3; ¯Cyclin D1, CDK4, Bcl-2; ↑p21, Bax; |
(Lin et al. 2015) [17] |
|||
|
|
Ursolic acid |
HT29 |
¯Viabilities; ↑apoptosis population; ¯p-EGFR, p-ERK1/2, p-p38, and p-JNK; ¯Bcl-2, Bcl-xl; ↑caspase 3 and 9 activity; |
(Shan et al. 2009) [62] |
|||
|
|
Ursolic acid |
SW-480, LoVo |
¯Viabilities; ¯clone formation; ¯migration ability; ¯MMP9; CDH1 expression; ¯p-Akt, p-ERK; ¯COX-2, PGE2; ↑NF-κB and p300 translocation; ↑apoptosis population; ↑PARP; ↑caspase 3 and 9 cleavage; ↑cytochrome c release; |
(Wang, Liu, et al. 2013) [36] |
|||
|
P. notoginseng |
Panaxadiol |
HCT-116 |
¯Viabilities; ↑apoptosis population; ↑G1 population; |
(Li et al. 2009) [20] |
|||
|
|
N-butanol fraction- |
HCT-116 |
¯Viabilities; ↑apoptosis population; |
(Wang et al. 2007) [25] |
|||
|
|
N-butanol fraction |
SW-480 |
¯Viabilities; ¯DNA synthesis; ↑apoptosis population; ↑S, G2 phase population; |
(Wang, Xie, et al. 2009) [18] |
|||
|
P. quinquefolius |
Steamed extracts |
HCT-116, SW-480 |
↑Apoptosis population; ¯MMP; ROS; ↑NF-κB activation; |
(Li et al. 2010) [63] |
|||
|
|
Ginseng extracts |
HCT-116 |
¯Viabilities; affect ephrin receptor pathway; |
(Luo et al. 2008) [64] |
|||
|
|
Ginsenoside Rh2 |
HCT-116, SW-480 |
¯Viabilities; ↑apoptosis population; ↑ROS; NF-κB activation; ↑Bax, Bad; ¯Bcl-2, Bcl-XL; ↑cytoplasmic vacuoles; |
(Li et al. 2011) [65] |
|||
|
|
Ginsenoside Rg3 |
SW-480 |
¯Migration ability; ¯NF-κB activity; ¯MMP-9, COX-2; |
(Song et al. 2015) [66] |
|||
|
|
Ginsenoside Rg3 |
HCT-116 |
¯Viabilities; ¯colony formation; ¯β-catenin nuclear translocation; ¯PNCA; |
(He et al. 2011) [22] |
|||
|
S. miltiorrhiza |
Cryptotanshinone |
SW-480, HCT-116, LoVo |
↓Viabilities; ↑apoptosis population; ↓anchorage independent growth; ↑caspase cleavage; ↓Bcl-2; ↓p-Stat3; ↓pEGFR; ↑p21 levels; ↓CyclinD1, survivin; |
(Li, Saud, et al. 2015) [24] |
|||
|
|
Tanshinone IIA |
Colo-205 |
↓Viabilities, ↑apoptosis population; ↑p53, p21; ↑cytochrome c release; ↑Fas; ↑caspase 8 and 3 cleavage; |
(Su et al. 2008) [58] |
|||
|
Z. officinale |
Ethanol extracts |
HCT-116, HT-29 |
↓Cell viability; ↑apoptosis population; ↑G1 phase population; |
(Abdullah et al. 2010) [27] |
|||
|
|
6-gingerol |
LoVo |
¯Viabilities; ¯cyclin A, B1, CDK1; ↑p27, p21; ↑ROS, p-p53; ↑G2 phase population; |
(Lin, Lin, and Tsay 2012) [29] |
|||
|
|
6-Gingerol |
HCT-116, HT-29 |
↓Viabilities; ↑apoptosis population; ↑G1 phase population; ↓cyclin D1; ↑nonsteroidal anti-inflammatory drug-activated gene-1; ↓β-catenin translocation; ↑PKCε, GSK-3β activation; |
(Lee, Cekanova, and Baek 2008) [28] |
|||
Table 2: In vivo study of Chinese herbal medicine for CRC treatment.
|
Species |
Extract/compound |
In vivo model |
Pharmacodynamic indicators |
Refs |
|
C. chinensis |
Berberine |
SW-620 and LoVo xenograft and lung metastasis |
↓Tumor volume; ↓tumor foci number; |
(Liu et al. 2015) [12] |
|
|
Berberine |
DMH and DSS-induced tumor |
↓Tumor number; ↓tumor incidence; |
(Wu et al. 2012) [67] |
|
|
Berberine |
APC Min/+ mice; |
¯Polyps number and size; ↓cyclin D1, c-Myc; |
(Zhang et al. 2013) [34] |
|
|
Berberine |
AOM and DSS-induced tumor |
↓Tumor number and size; ↓Ki-67, COX-2; ↑AMPK; ↓p-mTOR, p-p65; ↑caspase 3 cleavage; |
(Li, Hua, et al. 2015) [68] |
|
C. longa |
Curcumin |
C-26 xenograft; zebrafish; |
¯Vascular formation; ¯tumor volume; |
(Gou et al. 2011) [69] |
|
G. lucidum |
Triterpenes |
HT-29 xenograft |
↓Tumor volume; ↓Ki-67; ↑Beclin-1, LC-3; |
(Thyagarajan et al. 2010) [70] |
|
H. diffusa |
Ethanol extract |
HT-29 xenograft |
↓Tumor volume; ↓intratumoral microvessel density; ↓SHH, PTCH-1, SMO, Gli-1; ¯VEGF-A, VEGFR2; |
(Lin et al. 2013) [71] |
|
|
Ethanol extract |
HT-29 xenograft |
↓Tumor volume and weight; ↑apoptosis population; ↓PCNA; ↓p-Stat3; ↑Bax, p21; ↓Bcl-2, cyclin D1, CDK4; |
(Cai et al. 2012) [72] |
|
|
Aqueous extract |
HCT-116 xenograft |
↓Tumor volume and weight; ↑p-AMPK ;↓mTORC1; ↑p53 activation; |
(Lu et al. 2016) [38] |
|
P. quinquefolius |
protopanaxadiol |
HCT-116 xenograft |
↓Tumor size; ↓PCNA; |
(Gao et al. 2013) [12] |
|
R. Astragali |
Astragalus saponins |
HT-29 xenograft |
↓Tumor volume; ↓p-Akt, p-mTOR, VEGF, VEGFR1, VEGFR2; ↓COX-2; |
(Law et al. 2012) [73] |
|
|
Astragalus saponins |
HT-29 xenograft |
↓Tumor volume and weight; ↓PCNA; |
(Tin et al. [39] 2007) |
|
|
Astragalus saponins |
HCT-116 xenograft |
↓Tumor volume and weight; ↓PCNA; ↓VEGF; |
(Auyeung, Law, and Ko 2014) [41] |
|
S. miltiorrhiza |
Tanshinone IIA |
Colo-205 xenograft; |
↓Tumor volume; ↓P-glycoprotein, LC-3 II; ↓VEGF, NF-κB p65, and MMP 7; |
(Su 2012) [74] |
|
S. baicalensis |
Baicalein |
AOM and DSS-induced tumor |
↓Tumor number; |
(Kim et al. 2013) [32] |
|
|
Baicalein |
HCT-116 xenograft |
↓Tumor volume; |
(Wang et al. 2015) |
|
|
Baicalin |
HCT-116 xenograft |
↓Tumor volume; ↑survival rate; |
(Yang et al. 2013) [75] |
|
Z. officinale |
6-gingerol |
HCT-116 xenograft |
↓ tumor volume; ↓ leukotriene B4; ↓leukotriene A4 hydrolase; |
(Jeong et al. 2009) [76] |
References
- Fleming M, Ravula S, Tatishchev SF, Wang HL (2012) Colorectal carcinoma: pathologic aspects. J Gastrointest Oncol 3: 153-173. [Crossref]
- Su CC (2012) Tanshinone IIA potentiates the efficacy of 5-FU in Colo205 colon cancer cells in vivo through downregulation of P-gp and LC3-II. Exp Ther Med 3: 555-559.
- Sobin LH, Fleming ID (1997) TNM classification of malignant tumors, fifth edition (1997). Union Internationale Contre le Cancer and the American Joint Committee on Cancer. Cancer 80: 1803-1804. [Crossref]
- Bettington M, Walker N, Clouston A, Brown I, Leggett B et al. (2013) The serrated pathway to colorectal carcinoma: current concepts and challenges. Histopathology 62: 367-386. [Crossref]
- Fearon ER (2011) Molecular genetics of colorectal cancer. Annu Rev Pathol 6: 479-507. [Crossref]
- Cancer Genome Atlas, Network (2012) Comprehensive molecular characterization of human colon and rectal cancer. Nature 487: 330-337. [Crossref]
- Ling Y (2013) Traditional Chinese medicine in the treatment of symptoms in patients with advanced cancer. Ann Palliat Med 2: 141-152. [Crossref]
- Tanaka S, Haruma K, Kunihiro M, Nagata S, Kitadai Y et al. (2004) Effects of aged garlic extract (AGE) on colorectal adenomas: a double-blinded study. Hiroshima J Med Sci 53: 39-45. [Crossref]
- Hsiao WL, Liu L (2010) The role of traditional Chinese herbal medicines in cancer therapy-from TCM theory to mechanistic insights. Planta Med 76: 1118-1131. [Crossref]
- Li XK, Motwani M, Tong W, Bornmann W, Schwartz GK (2000) Huanglian, a Chinese herbal extract, inhibits cell growth by suppressing the expression of cyclin B1 and inhibiting CDC2 kinase activity in human cancer cells. Mol Pharmacol 58: 1287-1293. [Crossref]
- Kim YJ, Kang SA, Hong MS, Park HJ, Kim MJ et al. (2004) Coptidis rhizoma induces apoptosis in human colorectal cancer cells SNU-C4. Am J Chin Med 32: 873-882. [Crossref]
- Liu X, Ji Q, Ye N, Sui H, Zhou L et al. (2015) Berberine inhibits invasion and metastasis of colorectal cancer cells via COX-2/PGE2 mediated JAK2/STAT3 signaling pathway. PLoS One 10: e0123478. [Crossref]
- Lee YK, Park SY , Kim YM, Park OJ (2009) Regulatory effect of the AMPK-COX-2 signaling pathway in curcumin-induced apoptosis in HT-29 colon cancer cells. Ann N Y Acad Sci 1171: 489-494. [Crossref]
- Su CC, Lin JG, Li M, Chung JG, Yang JS et al. (2006) Curcumin-induced apoptosis of human colon cancer colo 205 cells through the production of ROS, Ca2+ and the activation of caspase-3. Anticancer Rese 26: 4379-4389. [Crossref]
- Guo LD, Chen XJ, Hu YH, Yu ZJ, Wang D, Liu JZ (2013) Curcumin inhibits proliferation and induces apoptosis of human colorectal cancer cells by activating the mitochondria apoptotic pathway. Phytother Res 27: 422-430. [Crossref]
- Li Q, Wang X, Shen A, Zhang Y, Chen Y et al. (2015) Hedyotis diffusa Willd overcomes 5-fluorouracil resistance in human colorectal cancer HCT-8/5-FU cells by downregulating the expression of P-glycoprotein and ATP-binding casette subfamily G member 2. Exp Ther Med 10: 1845-1850. [Crossref]
- Lin J, Li Q, Chen H, Lin H, Lai Z et al. (2015) Hedyotis diffusa Willd. extract suppresses proliferation and induces apoptosis via IL-6-inducible STAT3 pathway inactivation in human colorectal cancer cells. Oncol Lett 9: 1962-1970. [Crossref]
- Wang CZ, Zhang CF, Chen LN, Anderson S, Lu F et al. (2015) Colon cancer chemopreventive effects of baicalein, an active enteric microbiome metabolite from baicalin. Int J Oncol 47: 1749-1758. [Crossref]
- Wang CZ, Xie JT, Fishbein A, Aung HH, He H et al. (2009) Antiproliferative effects of different plant parts of Panax notoginseng on SW480 human colorectal cancer cells. Phytother Res 23: 6-13. [Crossref]
- Li XL, Wang CZ, Mehendale SR, Sun S, Wang Q et al. (2009) Panaxadiol, a purified ginseng component, enhances the anti-cancer effects of 5-fluorouracil in human colorectal cancer cells. Cancer Chemother Pharmacol 64: 1097-1104. [Crossref]
- Junmin S, Hongxiang L, Zhen L, Chao Y, Chaojie W (2015) Ginsenoside Rg3 inhibits colon cancer cell migration by suppressing nuclear factor kappa B activity. J Tradit Chin Med 35: 440-444. [Crossref]
- He BC, Gao JL, Luo XJ, Luo J, Shen J et al. (2011) Ginsenoside Rg3 inhibits colorectal tumor growth through the down-regulation of Wnt/beta-catenin signaling. Int J Oncol 38: 437-445. [Crossref]
- Su CC, Chen GW, Lin JG, Wu LT, Chung JG (2006) Curcumin inhibits cell migration of human colon cancer colo 205 cells through the inhibition of nuclear factor kappa B/p65 and down-regulates cyclooxygenase-2 and matrix metalloproteinase-2 expressions. Anticancer Res 26: 1281-1288. [Crossref]
- Li W, Saud SM, Young MR, Colburn NH, Hua B (2015) Cryptotanshinone, a Stat3 inhibitor, suppresses colorectal cancer proliferation and growth in vitro. Mol Cell Biochem 406: 63-73. [Crossref]
- Wang CZ, Luo X, Zhang B, Song WX, Ni M et al. (2007) Notoginseng enhances anti-cancer effect of 5-fluorouracil on human colorectal cancer cells. Cancer Chemother Pharmacol 60: 69-79. [Crossref]
- He LC, Lu N, Dai QS, Zhao Y, Zhao L et al. (2013) Wogonin induced G1 cell cycle arrest by regulating Wnt/beta-catenin signaling pathway and inactivating CDK8 in human colorectal cancer carcinoma cells. Toxicology 312: 36-47. [Crossref]
- Shailah A, Abidin SAZ, Murad NA, Makpol S, Wan Ngah WZ et al. (2010) Ginger extract (Zingiber officinale) triggers apoptosis and G0/G1 cells arrest in HCT 116 and HT 29 colon cancer cell lines. AJBR 4: 134-142.
- Lee SH, Cekanova M, Baek SJ (2008) Multiple mechanisms are involved in 6-gingerol-induced cell growth arrest and apoptosis in human colorectal cancer cells. Mol Carcinog 47: 197-208. [Crossref]
- Lin CB, Lin CC, Tsay GJ (2012) 6-Gingerol inhibits growth of colon cancer cell LoVo via induction of G2/M arrest. Evid Based Complement Alternat Med 2012: 326096. [Crossref]
- Xie JT, Wang CZ, Wicks S, Yin JJ, Kong J et al. (2006) Ganoderma lucidum extract inhibits proliferation of SW 480 human colorectal cancer cells. Exp Oncol 28: 25-29. [Crossref]
- Li Q, Wang X, Shen A, Zhang Y, Chen Y et al. (2015) Hedyotis diffusa Willd overcomes 5-fluorouracil resistance in human colorectal cancer HCT-8/5-FU cells by downregulating the expression of P-glycoprotein and ATP-binding casette subfamily G member 2. Exp Ther Med 10: 1845-1850. [Crossref]
- Kim DH, Hossain MA, Kang YJ, Jang JY, Lee YJ et al. (2013) Baicalein, an active component of Scutellaria baicalensis Georgi, induces apoptosis in human colon cancer cells and prevents AOM/DSS-induced colon cancer in mice. Int J Oncol 43: 1652-1658. [Crossref]
- Wang CZ, Zhang CF, Chen LN, Anderson S, Lu F et al. (2015) Colon cancer chemopreventive effects of baicalein, an active enteric microbiome metabolite from baicalin. Int J Oncol 47: 1749-1758. [Crossref]
- Zhong LLD, Chen HY, Cho WCS, Meng XM, Tong Y (2012) The efficacy of Chinese herbal medicine as an adjunctive therapy for colorectal cancer: a systematic review and meta-analysis. Complement Ther Med 20: 240-252. [Crossref]
- Yang YF, Ge JZ, Wu Y, Xu Y, Liang BY et al. (2008) Cohort study on the effect of a combined treatment of traditional Chinese medicine and western medicine on the relapse and metastasis of 222 patients with stage II and III colorectal cancer after radical operation. Chin J Integr Med 14: 251-256. [Crossref]
- Wang J, Liu L, Qiu H, Zhang X, Guo W et al. (2013) Ursolic acid simultaneously targets multiple signaling pathways to suppress proliferation and induce apoptosis in colon cancer cells. PloS One 8: e63872. [Crossref]
- Cai Q, Lin J, Wei L, Zhang L, Wang L et al. (2012) Hedyotis diffusa Willd inhibits colorectal cancer growth in vivo via inhibition of STAT3 signaling pathway. Int J Mol Sci 13: 6117-6128. [Crossref]
- Lu PH, Chen MB, Ji C, Li WT, Wei MX et al. (2016) Aqueous Oldenlandia diffusa extracts inhibits colorectal cancer cells via activating AMP-activated protein kinase signalings. Oncotarget 7: 45889-45900. [Crossref]
- Wang CZ, Calway TD, Wen XD, Smith J, Yu CH et al. (2013) Hydrophobic flavonoids from Scutellaria baicalensis induce colorectal cancer cell apoptosis through a mitochondrial-mediated pathway. Int J Oncol 42: 1018-1026. [Crossref]
- Law PC, Auyeung KK, Chan LY, Ko JK (2012) Astragalus saponins downregulate vascular endothelial growth factor under cobalt chloride-stimulated hypoxia in colon cancer cells. BMC Complement Altern Med 12: 160. [Crossref]
- Auyeung KKW, Law PC, Ko JKS (2014) Combined therapeutic effects of Vinblastine and Astragalus saponins in human colon cancer cells and tumor xenograft via inhibition of tumor growth and proangiogenic factors. Nutri Cancer Int J 66: 662-674. [Crossref]
- McCulloch M, Broffman M, van der Laan M, Hubbard A, Kushi L et al. (2011) Colon cancer survival with herbal medicine and vitamins combined with standard therapy in a whole-systems approach: ten-year follow-up data analyzed with marginal structural models and propensity score methods. Integr Cancer Ther 10: 240-259. [Crossref]
- Zhang JF, Cao HL, Zhang B, Cao HW, Xu XQ et al. (2013) Berberine potently attenuates intestinal polyps growth in ApcMin mice and familial adenomatous polyposis patients through inhibition of Wnt signalling. J Cell Mol Med 17: 1484-1493. [Crossref]
- Teas J, Pino S, Critchley A, Braverman LE (2004) Variability of iodine content in common commercially available edible seaweeds. Thyroid 14: 836-841. [Crossref]
- Farrell MP, Kummar S (2003) Phase I/IIA randomized study of PHY906, a novel herbal agent, as a modulator of chemotherapy in patients with advanced colorectal cancer. Clin Colorectal Cancer 2: 253-256. [Crossref]
- Zhou L, Shi P, Chen NH, Zhong JJ (2011) Ganoderic acid Me induces apoptosis through mitochondria dysfunctions in human colon carcinoma cells. Process Biochemistry 46: 219-225.
- Jayaprakasha GK, Rao LJM (2011) Chemistry, Biogenesis, and Biological Activities of Cinnamomum zeylanicum. Crit Rev Food Science Nutr 51: 547-562. [Crossref]
- Gruenwald J, Freder J, Armbruester N (2010) Cinnamon and Health. Crit Rev Food Sci Nutr 50: 822-834. [Crossref]
- Qin BL, Nagasaki M, Ren M, Bajotto G, Oshida Y et al. (2003) Cinnamon extract (traditional herb) potentiates in vivo insulin-regulated glucose utilization via enhancing insulin signaling in rats. Diabetes Res Clin Pract 62: 139-148. [Crossref]
- Abraham K, Wohrlin F, Lindtner O, Heinemeyer G, Lampen A (2010) Toxicology and risk assessment of coumarin: Focus on human data. Mol Nutr Food Res 54: 228-239. [Crossref]
- Ranasinghe P, Jayawardana R, Galappaththy P, Constantine GR, Gunawardana ND et al. (2012) Efficacy and safety of 'true' cinnamon (Cinnamomum zeylanicum) as a pharmaceutical agent in diabetes: a systematic review and meta-analysis. Diabet Med 29: 1480-1492. [Crossref]
- Thyagarajan A, Jedinak A, Nguyen H, Terry C, Baldridge LA et al. (2010) Triterpenes from Ganoderma Lucidum induce autophagy in colon cancer through the inhibition of p38 mitogen-activated kinase (p38 MAPK). Nutr Cancer 62: 630-640. [Crossref]
- Puranik AS, Halade G, Kumar S, Mogre R, Apte K et al. (2011) Cassia auriculata: Aspects of Safety Pharmacology and Drug Interaction. Evid Based Complement Alternat Med 2011: 915240.
- Teas J, Pino S, Critchley A, Braverman LE (2004) Variability of iodine content in common commercially available edible seaweeds. Thyroid 14: 836-841. [Crossref]
- Madabushi R, Frank B, Drewelow B, Derendorf H, Butterweck V (2006) Hyperforin in St. John's wort drug interactions. Eur J Clin Pharmacol 62: 225-233. [Crossref]
- Kim YJ, Kang SA, Hong MS, Park HJ, Kim MJ et al. (2004) Coptidis rhizoma induces apoptosis in human colorectal cancer cells SNU-C4. Am J Chin Med 32: 873-882. [Crossref]
- Du BY, Jiang LP, Xia Q, Zhong LF (2006) Synergistic inhibitory effects of curcumin and 5-fluorouracil on the growth of the human colon cancer cell line HT-29. Chemotherapy 52: 23-28. [Crossref]
- Su CC, Chen GW, Lin JG, Wu LT, Chung JG (2006) Curcumin inhibits cell migration of human colon cancer colo 205 cells through the inhibition of nuclear factor kappa B/p65 and down-regulates cyclooxygenase-2 and matrix metalloproteinase-2 expressions. Anticancer Res 26: 1281-1288. [Crossref]
- Chen A, Xu J, Johnson AC (2006) Curcumin inhibits human colon cancer cell growth by suppressing gene expression of epidermal growth factor receptor through reducing the activity of the transcription factor Egr-1. Oncogene 25: 278-287. [Crossref]
- Goel A, Boland CR, Chauhan DP (2001) Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett 172: 111-118. [Crossref]
- Lin J, Chen Y, Wei L, Chen X, Xu W et al. (2010) Hedyotis Diffusa Willd extract induces apoptosis via activation of the mitochondrion-dependent pathway in human colon carcinoma cells. Int J Oncol 37: 1331-1338.
- Shan JZ, Xuan YY, Zheng S, Dong Q, Zhang SZ (2009) Ursolic acid inhibits proliferation and induces apoptosis of HT-29 colon cancer cells by inhibiting the EGFR/MAPK pathway. J Zhejiang Univ Sci B 10: 668-674. [Crossref]
- Li BH, Wang CZ, He TC, Yuan CS, Du W (2010) Antioxidants potentiate American ginseng-induced killing of colorectal cancer cells. Cancer Lett 289: 62-70. [Crossref]
- Luo XJ, Wang CZ, Chen J, Song WX, Luo JY et al. (2008) Characterization of gene expression regulated by American ginseng and ginsenoside Rg3 in human colorectal cancer cells. Int J Oncol 32: 975-983. [Crossref]
- Li BH, Zhao JO, Wang CZ, Searle J, He TC et al. (2011) Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett 301: 185-192. [Crossref]
- Stewart Bernard W, Chris Wild, International Agency for Research on Cancer, World Health Organization (2014) World cancer report 2014. Lyon, France; Geneva, Switzerland: International Agency for Research on Cancer; WHO Press.
- Wu K, Yang Q, Mu Y, Zhou L, Liu Y et al. (2012) Berberine inhibits the proliferation of colon cancer cells by inactivating Wnt/beta-catenin signaling. Int J Oncol 41: 292-298. [Crossref]
- Li WD, Hua BJ, Saud SM, Lin HS, Hou W et al. (2015) Berberine regulates AMP-activated protein kinase signaling pathways and inhibits colon tumorigenesis in mice. Mol Carcinog 54: 1096-1109. [Crossref]
- Gou ML, Men K, Shi HS, Xiang ML, Zhang J et al. (2011) Curcumin-loaded biodegradable polymeric micelles for colon cancer therapy in vitro and in vivo. Nanoscale 3: 1558-1567. [Crossref]
- Thyagarajan A, Jedinak A, Nguyen H, Terry C, Baldridge LA et al. (2010) Triterpenes from Ganoderma Lucidum induce autophagy in colon cancer through the inhibition of p38 mitogen-activated kinase (p38 MAPK). Nutr Cancer 62: 630-640. [Crossref]
- Lin JM, Wei LH, Shen AL, Cai QY, Xu W et al. (2013) Hedyotis diffusa Willd extract suppresses Sonic hedgehog signaling leading to the inhibition of colorectal cancer angiogenesis. Int J Oncol 42: 651-656. [Crossref]
- Cai Q, Lin J, Wei L, Zhang L, Wang L et al. (2012) Hedyotis diffusa Willd inhibits colorectal cancer growth in vivo via inhibition of STAT3 signaling pathway. Int J Mol Sci 13: 6117-6128. [Crossref]
- Gao JL, Lv GY, He BC, Zhang BQ, Zhang HY et al. (2013) Ginseng saponin metabolite 20(S)-protopanaxadiol inhibits tumor growth by targeting multiple cancer signaling pathways. Oncol Rep 30: 292-298. [Crossref]
- Su CC, Chen GW, Kang JC, Chan MH (2008) Growth inhibition and apoptosis induction by tanshinone IIA in human colon adenocarcinoma cells. Planta Med 74: 1357-1362. [Crossref]
- Yang BL, Chen HJ, Chen YG, Gu YF, Zhang SP et al. (2013) Inhibitory effects of baicalin on orthotopic xenografts of colorectal cancer cells that are deficient in a mismatch repair gene in nude mice. Int J Colorectal Dis 28: 547-553. [Crossref]
- Jeong CH, Bode AM, Pugliese A, Cho YY, Kim HG et al. (2009) [6]-Gingerol suppresses colon cancer growth by targeting leukotriene A4 hydrolase. Cancer Res 69: 5584-5591. [Crossref]